The Impact of Seasonal Environments in a Tropical Savanna Climate on Forking, Leaf Area Index, and Biomass of Cassava Genotypes


 ,
,
Abstract
:1. Introduction
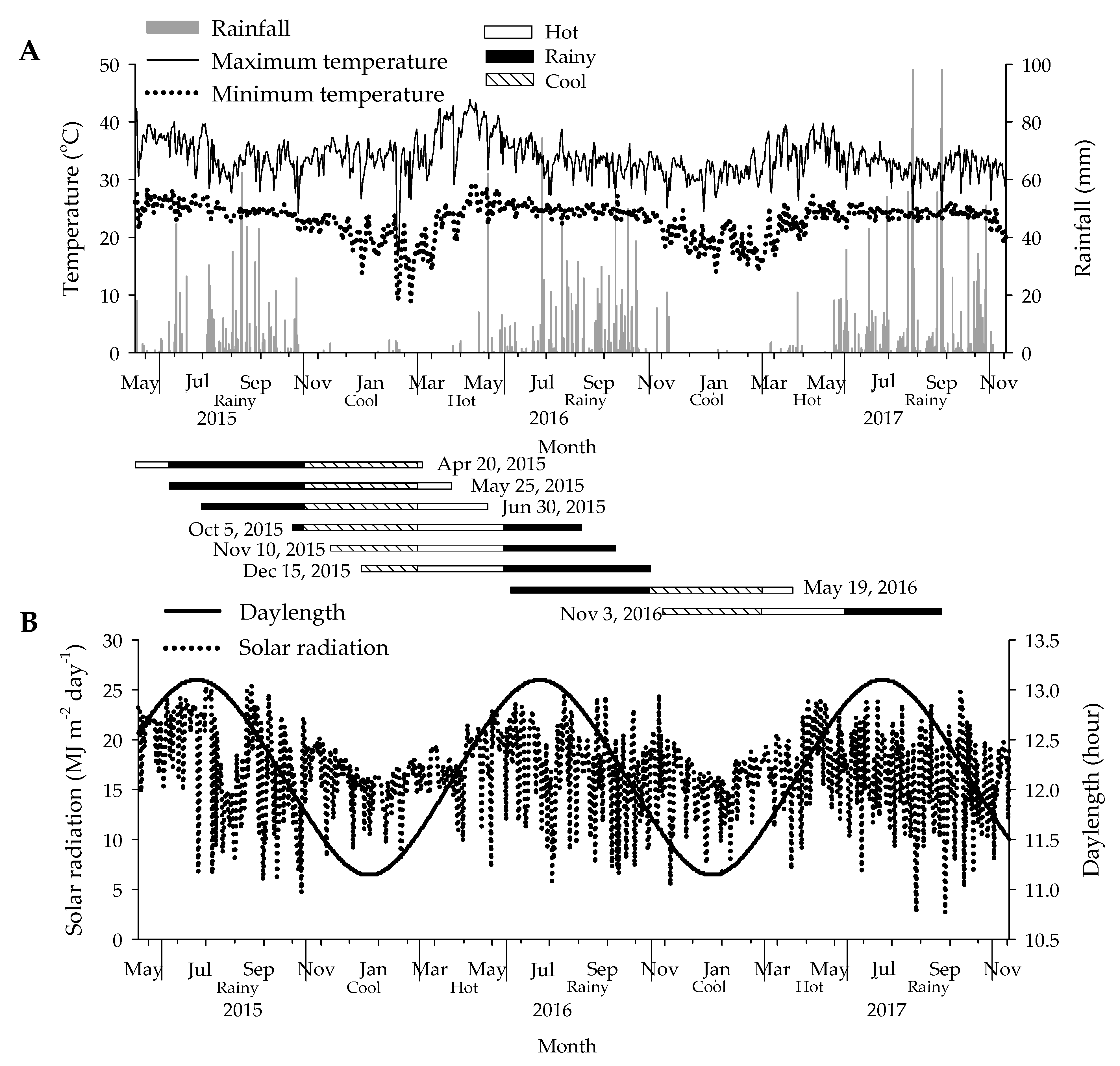
2. Materials and Methods
3. Results and Discussion
3.1. Combined Analysis
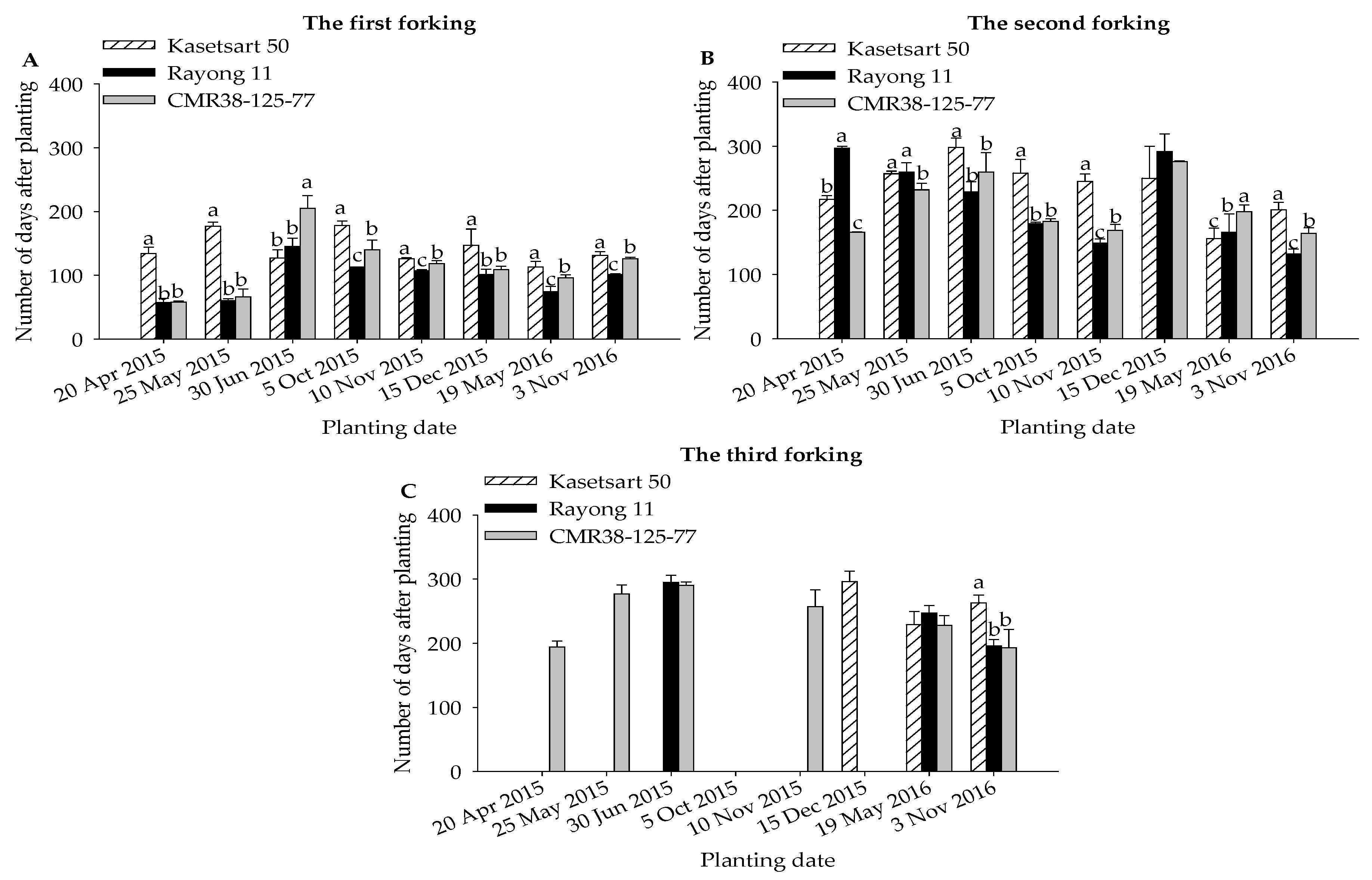
3.2. Forking Dates, Leaf Performances, and Final Harvest Data
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Howeler, R.H. Save and Grow: Cassava, A guide to sustainable production intensification; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- ASEAN Food Security Information System. ASEAN Agricultural Commodity Outlook; Office of Agricultural Economics, Ministry of Agriculture and Cooperatives: Bangkok, Thailand, 2015. [Google Scholar]
- Manrique, L.A. Leaf area development and dry matter production of cassava. Agron. J. 1990, 82, 887–891. [Google Scholar] [CrossRef]
- Keating, B.A.; Evenson, J.P.; Fukai, S. Environmental effects on growth and development of cassava (Manihot Esculenta Crantz.) I. Crop development. Field Crops Res. 1982, 5, 271–281. [Google Scholar] [CrossRef]
- Alves, A.A.C. Cassava Botany and Physiology. In Cassava: Biology, Production and Utilization; Hillocks, R.J., Thresh, J.M., Bellotti, A.C., Eds.; CABI Publishing: New York, NY, USA, 2002; pp. 67–89. [Google Scholar] [CrossRef]
- Tan, S.L.; Cock, J.H. Branching habit as a yield determinant in cassava. Field Crops Res. 1979, 2, 281–289. [Google Scholar] [CrossRef]
- Irikura, V.; Cock, J.H.; Kawano, K. The physiological basis of genotype–temperature interactions in cassava. Field Crops Res. 1979, 2, 227–239. [Google Scholar] [CrossRef]
- Cock, J.H. Cassava. In The Physiology of Tropical Field Crops; Goldsworthy, P.R., Fisher, N.M., Eds.; Wiley: New York, NY, USA, 1984; pp. 529–549. [Google Scholar] [CrossRef]
- El-Sharkawy, M.A. Cassava biology and physiology. Plant Mol. Biol. 2003, 53, 621–641. [Google Scholar] [CrossRef]
- Fukai, S.; Alcoy, A.B.; Llamelo, A.B.; Patterson, R.D. Effects of solar radiation on growth of cassava (Manihot esculenta Crantz.). I. Canopy development and dry matter growth. Field Crops Res. 1984, 9, 347–360. [Google Scholar] [CrossRef]
- El-Sharkawy, M.A. International research on cassava photosynthesis, productivity, eco–physiology, and responses to environmental stresses in the tropics. Photosynthetica 2006, 44, 481–512. [Google Scholar] [CrossRef]
- Howeler, R.H. Cassava Research and Development in Asia: Exploring New Opportunities for an Ancient Crop. In Proceedings of the 7th Regional Workshop, Bangkok, Thailand, 28 October–1 November 2002; Centro Internacional de Agricultura Tropical (CIAT), Cassava Office for Asia: Bangkok, Thailand, 2007. [Google Scholar]
- Meteorological Department. The Climate of Thailand. Available online: https://www.tmd.go.th/en/archive/thailand_climate.pdf (accessed on 4 September 2018).
- Department of Agriculture. Good Agricultural Practices for Cassava; National Bureau of Agricultural Commodity and Food Standards Ministry of Agriculture and Cooperatives: Bangkok, Thailand, 2008.
- Phoncharoen, P.; Banterng, P.; Vorasoot, N.; Jogloy, S.; Theerakulpisut, P.; Hoogenboom, G. Growth rates and yields of cassava at different planting dates in a tropical savanna climate. Sci. Agric 2018, in press. [Google Scholar]
- Howeler, R.H. Cassava mineral nutrition and fertilization. In Cassava: Biology, Production and Utilization; Hillocks, R.J., Thresh, J.M., Bellotti, A.C., Eds.; CABI Publishing: New York, NY, USA, 2002; pp. 115–147. [Google Scholar] [CrossRef]
- Richards, L.A.; Weaver, L.R. Moisture retention by some irrigated soils as related to soil moisture tension. J. Agric. Res. 1944, 69, 215–235. [Google Scholar]
- Wholey, D.W.; Booth, R.H. A comparison of simple methods for estimating starch content of cassava roots. J. Sci. Food Agric. 1979, 30, 158–164. [Google Scholar] [CrossRef]
- Statistix, version 10; Analytical Software: Tallahassee, FL, USA, 2013.
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley and Sons: New York, NY, USA, 1984. [Google Scholar]
- Phuntupan, K.; Banterng, P. Physiological determinants of storage root yield in three cassava genotypes under different nitrogen supply. J. Agr. Sci. Cambridge. 2017, 155, 978–992. [Google Scholar] [CrossRef]
- Sawatraksa, N.; Banterng, P.; Jogloy, S.; Vorasoot, N.; Hoogenboom, G. Chlorophyll fluorescence and biomass of four cassava genotypes grown under rain–fed upper paddy field conditions in the tropics. J. Agron. Crop Sci. 2018, 204, 554–565. [Google Scholar] [CrossRef]
- Veltkamp, H.J. Growth, total dry matter yield and its partitioning in cassava at different daylengths. Agric. Univ. Wageningen Papers 1985, 85, 73–86. [Google Scholar]
- Connor, D.J.; Cock, J.H. Response of cassava to water shortage. II. Canopy dynamics. Field Crops Res. 1981, 4, 285–296. [Google Scholar] [CrossRef]
- Lahai, M.T.; George, J.B.; Ekanayake, I.J. Cassava (Manihot esculenta Crantz) growth indices, root yield and its components in upland and inland valley ecologies of Sierra Leone. J. Agron. Crop Sci. 1999, 182, 239–247. [Google Scholar] [CrossRef]
- Lebot, V. Tropical Root and Tuber Crops: Cassava, Sweet Potato, Yam and Aroids; CABI: Wallingford, UK, 2009. [Google Scholar]
- El-Sharkawy, M.A.; De Tafur, S.M. Comparative photosynthesis, growth, productivity, and nutrient use efficiency among tall– and short–stemmed rain–fed cassava cultivars. Photosynthetica 2010, 48, 173–188. [Google Scholar] [CrossRef]
- Lahai, M.T.; Ekanayake, I.J.; Koroma, J.P.C. Influence of canopy structure on yield of cassava cultivars at various toposequences of an inland valley agro ecosystem. J. Agric. Biotech. Sustain. Dev. 2013, 5, 36–47. [Google Scholar] [CrossRef]
- Ramanujam, T. Leaf density profile and efficiency in partitioning dry matter among high and low yielding cultivars of cassava (Manihot esculenta Crantz). Field Crops Res. 1985, 10, 291–303. [Google Scholar] [CrossRef]
- Sawatraksa, N.; Banterng, P.; Jogloy, S.; Vorasoot, N.; Hoogenboom, G. cassava growth analysis of production during the off–season of paddy rice. Crop Sci. 2018, in press. [Google Scholar] [CrossRef]
- Lemoine, R.; Camera, S.L.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.-L.; Laloi, M.; Coutos-Thévenot, P.; Maurousset, L.; et al. Source–to–sink transport of sugar and regulation by environmental factors. Front Plant Sci. 2013, 4, 1–21. [Google Scholar] [CrossRef]
- Kawano, K. Harvest index and evoluation of major food crop cultivars in the tropics. Euphytica 1990, 46, 195–202. [Google Scholar] [CrossRef]
- Ceballos, H.; Iglesias, C.A.; Pérez, J.C.; Dixon, A.G. Cassava breeding: Opportunities and challenges. Plant Mol. Biol. 2004, 56, 503–516. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Soil Characteristic | Planting Date | |||||||
|---|---|---|---|---|---|---|---|---|
| 20 April 2015 | 25 May 2015 | 30 June 2015 | 5 October 2015 | 10 November 2015 | 15 December 2015 | 19 May 2016 | 3 November 2016 | |
| pH | 7.30 | 6.27 | 5.04 | 5.29 | 5.64 | 5.39 | 7.25 | 7.37 |
| Cation exchange capacity (cmol kg−1) | 6.62 | 7.80 | 3.52 | 4.60 | 6.38 | 6.99 | 2.89 | 2.83 |
| Organic matter (g kg−1) | 3.71 | 2.70 | 2.86 | 4.79 | 3.08 | 2.74 | 4.76 | 5.14 |
| Total nitrogen (g kg−1) | 0.18 | 0.15 | 0.14 | 0.21 | 0.13 | 0.14 | 0.24 | 0.35 |
| Available phosphorus (mg kg−1) | 56.85 | 45.02 | 9.89 | 16.10 | 32.60 | 15.45 | 82.26 | 85.13 |
| Exchangeable potassium (mg kg−1) | 27.63 | 27.70 | 17.86 | 36.42 | 52.31 | 21.82 | 33.46 | 31.48 |
| Crop Trait | Source of Variation | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Planting Date (PD) | Replication/PD | Genotype (G) | GxPD | Pool Error | ||||||
| MS | %SS | MS | %SS | MS | %SS | MS | %SS | MS | %SS | |
| The first forking date a | 7524.5 | 36.7 ** | 95.9 | 1.6 | 18609.8 | 26.0 ** | 3327.3 | 32.5 ** | 95.0 | 3.2 |
| The second forking date a | 20300.7 | 54.0 ** | 431.8 | 3.9 | 7480.0 | 5.7 ** | 6042.7 | 32.1 ** | 232.8 | 4.2 |
| TDW at 300 DAP (t ha−1) | 248.7 | 39.4 ** | 12.9 | 7.0 | 156.0 | 10.6 ** | 60.4 | 28.7 ** | 8.8 | 14.3 |
| SRDW at 300 DAP (t ha−1) | 54.5 | 16.5 ** | 6.5 | 6.8 | 116.5 | 15.1 ** | 50.1 | 45.5 ** | 5.2 | 16.1 |
| HI at 300 DAP | 0.1 | 45.5 ** | 0.0 | 6.7 | 0.1 | 17.7 ** | 0.0 | 21.3 ** | 0.0 | 8.7 |
| Starch content at 300 DAP (%) | 147.0 | 59.3 ** | 2.3 | 3.2 | 44.2 | 7.6 ** | 13.6 | 16.5 ** | 3.2 | 13.4 |
| LAI at 90 DAP (cm2 cm−2) | 45.9 | 88.6 ** | 0.3 | 1.7 | 3.2 | 2.6 ** | 0.8 | 4.5 ** | 0.1 | 2.5 |
| LAI at 270 DAP (cm2 cm−2) | 17.6 | 53.7 ** | 0.2 | 2.6 | 17.7 | 23.2 ** | 1.1 | 9.9 ** | 0.3 | 10.5 |
| LDW at 90 DAP (t ha−1) | 21.5 | 93.8 ** | 0.1 | 2.0 | 0.4 | 0.7 ** | 0.1 | 1.7 ** | 0.0 | 1.8 |
| LDW at 270 DAP (t ha−1) | 2.5 | 52.6 ** | 0.0 | 3.2 | 2.5 | 22.7 ** | 0.2 | 10.1 ** | 0.1 | 11.4 |
| Genotype | Variable | Coefficient | t | Determination Coefficient (R2) |
|---|---|---|---|---|
| Kasetsart 50 | The first forking | |||
| Constant | 142.22 | 32.54 ** | 0.00 | |
| The second forking | ||||
| Constant | 78.14 | 0.23 ns | 0.34 | |
| Solar radiation | −36.87 | −2.36 ** | ||
| Maximum temperature | 22.74 | 3.19 ** | ||
| Rayong 11 | The first forking | |||
| Constant | 578.34 | 8.19 ** | 0.81 | |
| Solar radiation | −14.50 | −5.49 ** | ||
| Minimum temperature | 17.28 | 5.84 ** | ||
| Daylength | −51.38 | −4.63 ** | ||
| The second forking | ||||
| Constant | −1110.15 | −4.62 ** | 0.50 | |
| Solar radiation | 80.02 | 5.52 ** | ||
| CMR38–125–77 | The first forking | |||
| Constant | 1210.63 | 8.37 ** | 0.83 | |
| Solar radiation | −24.20 | −6.77 ** | ||
| Minimum temperature | 41.05 | 7.04 ** | ||
| Daylength | −121.53 | −5.92 ** | ||
| The second forking | ||||
| Constant | −293.64 | −1.46 ns | 0.35 | |
| Solar radiation | −70.24 | −3.74 ** | ||
| Daylength | 138.96 | 3.90 ** |
| Planting Date | Genotype | LAI ± SD (cm2 cm−2) | Leaf dry weight ± SD (t ha−1) | ||
|---|---|---|---|---|---|
| 90 DAP | 270 DAP | 90 DAP | 270 DAP | ||
| 20 April 2015 | Kasetsart 50 | 5.7 ± 0.92 b | 2.9 ± 0.79 ab | 4.0 ± 0.77 | 1.1 ± 0.28 ab |
| Rayong 9 | 5.6 ± 0.63 b | 2.2 ± 0.64 bc | 3.5 ± 0.64 | 0.9 ± 0.20 b | |
| Rayong 11 | 6.5 ± 0.56 ab | 3.3 ± 0.58 a | 4.2 ± 0.69 | 1.4 ± 0.07 a | |
| CMR38–125–77 | 7.3 ± 0.53 a | 1.7 ± 0.35 c | 4.4 ± 0.20 | 0.7 ± 0.21 b | |
| F | ** | * | ns | ** | |
| 25 May 2015 | Kasetsart 50 | 3.4 ± 0.30 b | 2.9 ± 0.44 b | 1.3 ± 0.12 | 1.5 ± 0.37 ab |
| Rayong 9 | 3.2 ± 0.15 b | 2.4 ± 0.46 b | 1.3 ± 0.16 | 1.2 ± 0.10 b | |
| Rayong 11 | 3.3 ± 0.55 b | 4.1 ± 0.97 a | 1.1 ± 0.16 | 2.0 ± 0.36 a | |
| CMR38–125–77 | 4.3 ± 0.69 a | 2.8 ± 0.65 b | 1.4 ± 0.11 | 1.3 ± 0.25 b | |
| F | * | * | ns | * | |
| 30 June 2015 | Kasetsart 50 | 1.6 ± 0.41 b | 2.6 ± 0.50 ab | 0.6 ± 0.11 b | 1.4 ± 0.23 a |
| Rayong 9 | 1.5 ± 0.31 b | 1.4 ± 0.14 c | 0.6 ± 0.10 b | 0.7 ± 0.08 b | |
| Rayong 11 | 2.1 ± 0.47 b | 3.8 ± 0.88 a | 0.8 ± 0.12 ab | 1.7 ± 0.37 a | |
| CMR38–125–77 | 3.2 ± 0.37 a | 1.6 ± 0.24 bc | 1.0 ± 0.14 a | 0.8 ± 0.12 b | |
| F | ** | ** | ** | ** | |
| 5 October 2015 | Kasetsart 50 | 1.7 ± 0.12 a | 4.0 ± 0.46 ab | 0.9 ± 0.05 a | 1.9 ± 0.09 a |
| Rayong 9 | 1.2 ± 0.05 b | 2.8 ± 0.49 b | 0.6 ± 0.04 b | 1.3 ± 0.12 b | |
| Rayong 11 | 1.5 ± 0.35 ab | 4.7 ± 0.75 a | 0.7 ± 0.18 ab | 1.8 ± 0.33 a | |
| CMR38–125–77 | 1.7 ± 0.23 a | 4.3 ± 0.64 a | 0.9 ± 0.09 a | 1.7 ± 0.23 ab | |
| F | * | ** | * | * | |
| 10 November 2015 | Kasetsart 50 | 1.7 ± 0.51 b | 5.5 ± 0.70 a | 0.9 ± 0.27 | 2.3 ± 0.31 a |
| Rayong 9 | 1.8 ± 0.04 b | 3.2 ± 0.30 b | 0.9 ± 0.10 | 1.5 ± 0.05 b | |
| Rayong 11 | 2.5 ± 0.12 a | 5.9 ± 0.37 a | 1.2 ± 0.05 | 2.3 ± 0.16 a | |
| CMR38–125–77 | 2.5 ± 0.61 a | 4.0 ± 0.41 b | 1.2 ± 0.30 | 1.7 ± 0.26 b | |
| F | * | ** | ns | ** | |
| 15 December 2015 | Kasetsart 50 | 1.3 ± 0.11 ab | 3.7 ± 0.61 ab | 0.7 ± 0.09 a | 1.7 ± 0.21 a |
| Rayong 9 | 1.1 ± 0.22 b | 2.9 ± 0.50 b | 0.5 ± 0.09 b | 1.2 ± 0.26 b | |
| Rayong 11 | 1.3 ± 0.10 ab | 4.6 ± 0.43 a | 0.7 ± 0.04 a | 1.9 ± 0.13 a | |
| CMR38–125–77 | 1.5 ± 0.18 a | 3.5 ± 0.54 b | 0.7 ± 0.10 a | 1.4 ± 0.19 ab | |
| F | * | * | ** | ** | |
| 19 May 2016 | Kasetsart 50 | 2.4 ± 0.38 a | 0.8 ± 0.14 | 1.1 ± 0.16 a | 0.5 ± 0.07 |
| Rayong 9 | 1.2 ± 0.47 b | 1.6 ± 0.40 | 0.5 ± 0.20 b | 0.9 ± 0.21 | |
| Rayong 11 | 1.3 ± 0.32 b | 1.6 ± 0.96 | 0.6 ± 0.14 b | 0.8 ± 0.44 | |
| CMR38–125–77 | 1.0 ± 0.43 b | 1.1 ± 0.36 | 0.4 ± 0.17 b | 0.6 ± 0.18 | |
| F | ** | ns | ** | ns | |
| 3 November 2016 | Kasetsart 50 | 1.2 ± 0.15 | 3.1 ± 0.36 bc | 0.6 ± 0.05 | 1.2 ± 0.13 b |
| Rayong 9 | 1.1 ± 0.18 | 2.0 ± 0.27 c | 0.5 ± 0.08 | 0.9 ± 0.22 c | |
| Rayong 11 | 1.2 ± 0.21 | 4.6 ± 0.09 a | 0.6 ± 0.09 | 1.5 ± 0.10 a | |
| CMR38–125–77 | 1.2 ± 0.12 | 3.4 ± 0.91 b | 0.6 ± 0.08 | 1.0 ± 0.09 bc | |
| F | ns | ** | ns | ** | |
| Planting Date | Genotype | Total crop dry weight ± SD (t ha−1) | Storage root dry weight ± SD (t ha−1) | HI ± SD | Starch content ± SD (%) |
|---|---|---|---|---|---|
| 20 April 2015 | Kasetsart 50 | 36.2 ± 3.95 ab | 11.6 ± 2.69 bc | 0.32 ± 0.05 c | 30.7 ± 1.25 a |
| Rayong 9 | 41.1 ± 3.03 a | 21.6 ± 2.04 a | 0.53 ± 0.03 a | 32.3 ± 0.70 a | |
| Rayong 11 | 33.9 ± 2.88 bc | 14.5 ± 1.54 b | 0.43 ± 0.02 b | 29.6 ± 1.46 ab | |
| CMR38–125–77 | 28.8 ± 2.60 c | 7.9 ± 1.69 c | 0.27 ± 0.04 c | 27.0 ± 1.44 b | |
| F | ** | ** | ** | ** | |
| 25 May 2015 | Kasetsart 50 | 27.0 ± 2.80 b | 10.7 ± 0.72 b | 0.40 ± 0.03 ab | 21.8 ± 1.01 b |
| Rayong 9 | 29.9 ± 4.09 b | 13.4 ± 2.21 b | 0.45 ± 0.02 ab | 28.8 ± 1.95 a | |
| Rayong 11 | 30.9 ± 1.52 ab | 11.4 ± 1.51 b | 0.37 ± 0.06 b | 27.5 ± 0.47 a | |
| CMR38–125–77 | 35.8 ± 1.76 a | 17.4 ± 0.43 a | 0.49 ± 0.04 a | 28.9 ± 2.37 a | |
| F | ** | ** | ** | ** | |
| 30 June 2015 | Kasetsart 50 | 20.9 ± 2.74 ab | 7.9 ± 1.04 b | 0.38 ± 0.02 c | 22.5 ± 1.42 b |
| Rayong 9 | 19.4 ± 2.65 b | 12.3 ± 1.24 a | 0.64 ± 0.03 a | 25.4 ± 1.01 a | |
| Rayong 11 | 23.2 ± 0.47 ab | 12.0 ± 1.39 a | 0.52 ± 0.70 b | 24.3 ± 0.65 a | |
| CMR38–125–77 | 25.4 ± 0.94 a | 14.8 ± 0.83 a | 0.58 ± 0.02 ab | 24.2 ± 0.74 ab | |
| F | ** | ** | ** | * | |
| 5 October 2015 | Kasetsart 50 | 26.4 ± 1.96 b | 12.8 ± 1.35 b | 0.48 ± 0.02 b | 21.3 ± 0.62 |
| Rayong 9 | 25.0 ± 1.62 bc | 13.6 ± 1.29 b | 0.54 ± 0.05 a | 22.7 ± 2.60 | |
| Rayong 11 | 24.2 ± 1.31 c | 10.8 ± 0.57 c | 0.45 ± 0.02 b | 22.7 ± 1.25 | |
| CMR38–125–77 | 29.3 ± 1.51 a | 15.8 ± 0.99 a | 0.54 ± 0.02 a | 23.7 ± 1.89 | |
| F | ** | ** | ** | ns | |
| 10 November 2015 | Kasetsart 50 | 32.6 ± 1.32 a | 15.6 ± 1.36 ab | 0.48 ± 0.04 b | 24.6 ± 1.92 |
| Rayong 9 | 30.0 ± 2.02 ab | 17.9 ± 1.89 a | 0.60 ± 0.03 a | 26.1 ± 0.59 | |
| Rayong 11 | 25.1 ± 3.87 b | 11.3 ± 2.34 b | 0.45 ± 0.04 b | 23.9 ± 1.80 | |
| CMR38–125–77 | 34.0 ± 1.86 a | 18.2 ± 1.54 a | 0.54 ± 0.03 ab | 24.2 ± 1.41 | |
| F | ** | ** | ** | ns | |
| 15 December 2015 | Kasetsart 50 | 29.9 ± 1.85 b | 13.9 ± 1.51 b | 0.47 ± 0.08 bc | 23.7 ± 1.80 |
| Rayong 9 | 27.0 ± 1.15 b | 14.0 ± 1.28 b | 0.52 ± 0.04 ab | 26.0 ± 2.70 | |
| Rayong 11 | 22.1 ± 1.53 c | 9.5 ± 0.96 c | 0.43 ± 0.04 c | 23.8 ± 2.02 | |
| CMR38–125–77 | 34.6 ± 3.36 a | 18.5 ± 2.80 a | 0.54 ± 0.10 a | 24.1 ± 1.92 | |
| F | ** | ** | * | ns | |
| 19 May 2016 | Kasetsart 50 | 23.1 ± 5.77 | 11.7 ± 4.87 | 0.49 ± 0.13 b | 29.5 ± 3.21 |
| Rayong 9 | 21.2 ± 4.37 | 15.2 ± 3.07 | 0.72 ± 0.03 a | 31.2 ± 1.30 | |
| Rayong 11 | 27.4 ± 3.47 | 18.2 ± 1.88 | 0.67 ± 0.02 a | 31.2 ± 1.48 | |
| CMR38–125–77 | 28.0 ± 4.38 | 17.0 ± 3.16 | 0.61 ± 0.04 ab | 33.0 ± 0.71 | |
| F | ns | ns | ** | ns | |
| 3 November 2016 | Kasetsart 50 | 34.1 ± 4.94 a | 21.6 ± 4.86 a | 0.63 ± 0.05 a | 21.0 ± 1.65 b |
| Rayong 9 | 23.5 ± 5.45 b | 14.8 ± 4.56 b | 0.62 ± 0.09 a | 23.3 ± 4.06 ab | |
| Rayong 11 | 22.4 ± 4.74 b | 11.5 ± 4.09 b | 0.50 ± 0.08 b | 27.6 ± 0.78 a | |
| CMR38–125–77 | 33.5 ± 3.91 a | 22.8 ± 2.81 a | 0.68 ± 0.04 a | 28.2 ± 0.98 a | |
| F | * | * | * | ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phoncharoen, P.; Banterng, P.; Vorasoot, N.; Jogloy, S.; Theerakulpisut, P.; Hoogenboom, G. The Impact of Seasonal Environments in a Tropical Savanna Climate on Forking, Leaf Area Index, and Biomass of Cassava Genotypes. Agronomy 2019, 9, 19. https://doi.org/10.3390/agronomy9010019
Phoncharoen P, Banterng P, Vorasoot N, Jogloy S, Theerakulpisut P, Hoogenboom G. The Impact of Seasonal Environments in a Tropical Savanna Climate on Forking, Leaf Area Index, and Biomass of Cassava Genotypes. Agronomy. 2019; 9(1):19. https://doi.org/10.3390/agronomy9010019
Chicago/Turabian StylePhoncharoen, Phanupong, Poramate Banterng, Nimitr Vorasoot, Sanun Jogloy, Piyada Theerakulpisut, and Gerrit Hoogenboom. 2019. "The Impact of Seasonal Environments in a Tropical Savanna Climate on Forking, Leaf Area Index, and Biomass of Cassava Genotypes" Agronomy 9, no. 1: 19. https://doi.org/10.3390/agronomy9010019