Near-Isogenic Lines of Japonica Rice Revealed New QTLs for Cold Tolerance at Booting Stage



Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Traits
2.2. Extraction and Amplification of DNA
2.3. Construction of DNA Bulks and Molecular Marker Analysis
2.4. Mapping and QTL Analysis
3. Results
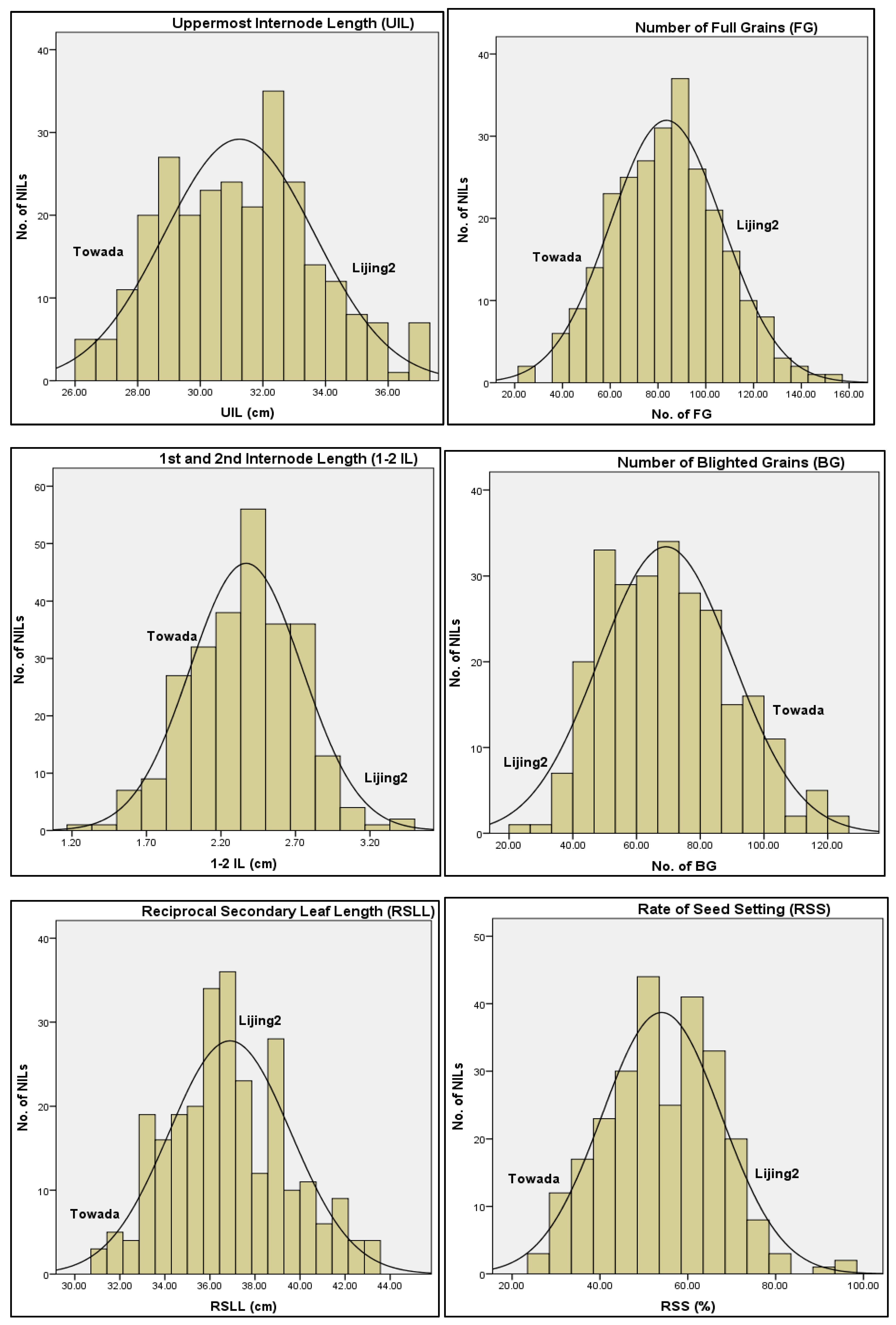
3.1. Growth Attributes and Cold Stress Tolerance
3.1.1. Correlations between Morphological Traits and RSS
3.1.2. Variation of Six Morphological Traits Correlated with Rice Cold tolerance at Booting Stage
3.2. Polymorphism of Molecular Markers (SSR)
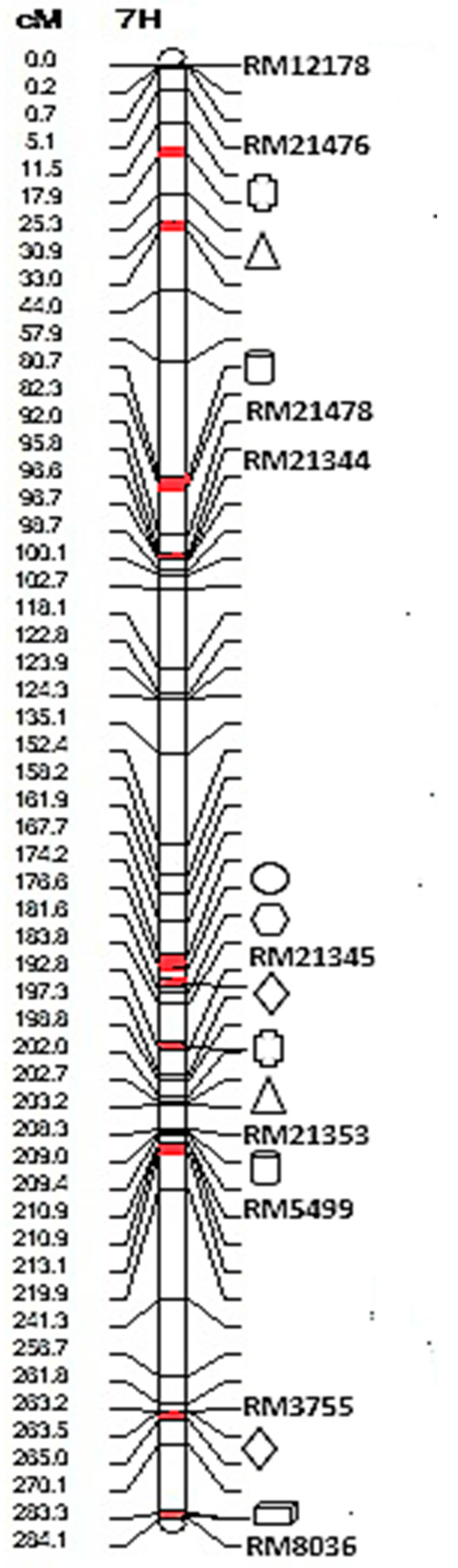
3.3. Linkage Mapping and Analysis of QTLs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wahid, A.; Gelani, S.; Ashraf, A.; Foolad, M.R. Heat Tolerance in Plants: An Overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Sims, L.; Pastor, J.; Lee, T.; Dewey, B. Nitrogen, phosphorus and light effects on growth and allocation of biomass and nutrients in wild rice. Oecologia 2012, 170, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Genisel, M.; Turk, H.; Erdal, S. Exogenous progesterone application protects chickpea seedlings against chilling-induced oxidative stress. Acta Physiol. Plant. 2013, 35, 241–251. [Google Scholar] [CrossRef]
- Zhu, J.; Dong, C.H.; Zhu, J.K. Interplay between cold-responsive gene regulation, metabolism and RNA processing during plant cold acclimation. Curr. Opin. Plant Biol. 2007, 10, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, Q.; Wang, S.; Hong, Y.; Wang, Z. Rice and cold stress: Methods for its evaluation and summary of cold tolerance-related quantitative trait loci. Rice 2014, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Kato, H.; Imai, R. Biochemical identification of the OsMKK6-OsMPK3 signalling pathway for chilling stress tolerance in rice. Biochem. J. 2012, 443, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Nagasuga, K.; Murai-Hatano, M.; Kuwagata, T. Effects of low root temperature on dry matter production and root water uptake in rice plants. Plant Prod. Sci. 2011, 14, 22–29. [Google Scholar] [CrossRef]
- Oliver, S.N.; Van Dongen, J.T.; Alfred, S.C.; Mamun, E.A.; Zhao, X.; Saini, H.S.; Fernandes, S.F.; Blanchard, C.L.; Sutton, B.G.; Geigenberger, P.; et al. Cold-induced repression of the rice anther-specific cell wall invertase gene OSINV4 is correlated with sucrose accumulation and pollen sterility. Plant Cell Environ. 2005, 28, 1534–1551. [Google Scholar] [CrossRef]
- Huang, X.; Kurata, N.; Wei, X.; Wang, Z.X. A map of rice genome variation reveals the origin of cultivated rice. Nature 2012, 490, 497–501. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.K.; Azhar, A.; Salam, I.; Ali, S.I.; Galani, S. Mitigating heat induced damages in rice (Oryza sativa L.) by hsp70 modulation. Pak. J. Bot. 2018, 50, 2181–2187. [Google Scholar]
- Sun, J.; Yang, L.; Wang, J.; Liu, H.; Zheng, H.; Xie, D.; Zhang, M.; Feng, M.; Jia, Y.; Zhao, H.; et al. Identification of a cold-tolerant locus in rice (Oryza sativa L.) using bulked segregant analysis with a next-generation sequencing strategy. Rice 2018, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.W.; Yang, S.M.; Cui, H.; Yang, X.J.; Xu, L.M.; Du, J.; Pu, X.Y.; Li, Z.C.; Cheng, Z.Q.; Huang, X.Q. QTLs of Cold Tolerance-related Traits at the Booting Stage for NIL-RILs in Rice Revealed by SSR. Genes Genom. 2009, 31, 143–154. [Google Scholar] [CrossRef]
- Lou, Q.J.; Chen, L.; Sun, Z.X.; Xing, Y.Z.; Li, J.; Xu, X.Y.; Mei, H.W.; Luo, L.J. A major QTL associated with cold tolerance at seedling stage in rice (Oryza sativa L.). Euphytica 2007, 158, 87–94. [Google Scholar] [CrossRef]
- Jena, K.K.; Kim, S.M.; Suh, J.P.; Yang, C.I.; Kim, Y.J. Identification of Cold-Tolerant Breeding Lines by Quantitative Trait Loci Associated with Cold Tolerance in Rice. Crop Sci. 2012, 51, 517–523. [Google Scholar] [CrossRef]
- Liu, F.X.; Sun, C.Q.; Tan, L.B.; Fu, Y.C.; Li, D.J.; Wang, X.K. Identification and mapping of quantitative trait loci controlling cold-tolerance of Chinese common wild rice (O. rufipogon Griff.) at booting to flowering stages. Chin. Sci. Bull. 2003, 48, 2068–2071. [Google Scholar] [CrossRef]
- Ye, C.; Fukai, S.; Godwin, I.; Reinke, R.; Snell, P.; Schiller, J.; Basnayake, J. Cold tolerance in rice varieties at different growth stages. Crop Pasture Sci. 2009, 60, 328–338. [Google Scholar] [CrossRef]
- Andaya, V.C.; Mackill, D.J. Mapping of QTLs associated with cold tolerance during the vegetative stage in rice. J. Exp. Bot. 2003, 54, 2579–2585. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Redoña, E.; Fukai, S.; Godwin, I.D.; Koh, H.; Reinke, R.; Zhou, Y.; Lambrides, C.; Jiang, W.; Snell, P. A QTL controlling low temperature induced spikelet sterility at booting stage in rice. Euphytica 2010, 176, 291–301. [Google Scholar] [CrossRef]
- Shimono, H.; Okada, M.; Kanda, E.; Arakawa, I. Low temperature-induced sterility in rice: Evidence for the effects of temperature before panicle initiation. Field Crops Res. 2007, 101, 221–231. [Google Scholar] [CrossRef]
- Farrell, T.C.; Fox, K.M.; Williams, R.L.; Fukai, S.; Lewin, L.G. Minimizing cold damage during reproductive development among temperate rice genotypes. II. Genotypic variation and flowering traits related to cold tolerance screening. Aust. J. Agric. Res. 2006, 57, 89–100. [Google Scholar] [CrossRef]
- Gunawardena, T.A.; Fukai, S.; Blamey, F.P.C. Low temperature induced spikelet sterility in rice. I. Nitrogen fertilisation and sensitive reproductive period. Aust. J. Agric. Res. 2003, 54, 937–946. [Google Scholar] [CrossRef]
- Ha, L.V.; Mitchell, J.H.; Fukai, S. Genotypic consistency for low temperature tolerance at the booting stage in rice grown under flooded and non-flooded conditions. Field Crops Res. 2018, 220, 19–26. [Google Scholar] [CrossRef]
- Xu, L.M.; Zhou, L.; Zeng, Y.W.; Wang, F.M.; Zhang, H.L.; Shen, S.Q.; Li, Z.C. Identification and mapping of quantitative trait loci for cold tolerance at the booting stage in a japonica rice nearisogenic line. Plant Sci. 2008, 174, 340–347. [Google Scholar] [CrossRef]
- Farrell, T.C.; Fox, K.M.; Williams, R.L.; Fukai, S. Genotypic variation for cold tolerance during reproductive development in rice: Screening with cold air and cold water. Field Crops Res. 2006, 98, 178–194. [Google Scholar] [CrossRef]
- Gunawardena, T.A.; Fukai, S.; Blamey, F.P.C. Low temperature induced spikelet sterility in rice. II. Effects of panicle and root temperatures. Aust. J. Agric. Res. 2003, 54, 947–956. [Google Scholar] [CrossRef]
- Cui, D.; Xu, C.Y.; Tang, C.F.; Yang, C.G.; Yu, T.Q. Genetic structure and association mapping of cold tolerance in improved japonica rice germplasm at the booting stage. Euphytica 2013, 193, 369–382. [Google Scholar] [CrossRef]
- Satake, T.; Shibata, M. Male-sterility caused by cooling treatment at the young microspore stage in rice plants. XXXI. Four components participating in fertilization. Jpn. J. Crop Sci. 1992, 61, 454–462. [Google Scholar] [CrossRef]
- Hu, H.H.; You, J.; Fang, Y.J.; Zhu, X.Y.; Qi, Z.Y.; Xiong, L.Z. Characterization of transcription factor gene SNAC2 conferring cold and salt tolerance in rice. Plant Mol. Biol. 2008, 67, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Andaya, V.C.; Tai, T.H. Fine mapping of the qCTS12 locus, a major QTL for seedling cold tolerance in rice. Theor. Appl. Genet. 2006, 113, 467–475. [Google Scholar] [CrossRef]
- Zhang, Q.; Jiang, N.; Wang, G.L.; Hong, Y.; Wang, Z. Advances in understanding cold sensing and the cold-responsive network in rice. Adv. Crop Sci. Tech. 2013, 1, 104. [Google Scholar]
- Zhou, L.; Zeng, Y.; Hu, G.; Pan, Y.; Yang, S.; You, A.; Zhang, H.; Li, J.; Li, Z. Characterization and identification of cold tolerant near-isogenic lines in rice. Breed. Sci. 2012, 62, 196–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Dai, C.C.; Cheng, Y.; Chen, T.; Jiao, D.M. Identification for cold tolerance at different growth stages in rice (Oryza sativa L.) and physiological mechanism of differential cold tolerance. Acta Agron. Sin. 2006, 32, 76–83. [Google Scholar]
- Kim, S.J.; Lee, S.C.; Hong, S.K.; An, K.; An, G.; Kim, S.R. Ectopic Expression of a Cold-Responsive OsAsr1 cDNA Gives Enhanced Cold Tolerance in Transgenic Rice Plants. Mol. Cells 2009, 27, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, J.; Wang, Y.Y.; Chen, Z.J.; Tu, S.S.; Feng, L.L.; Zhu, Y.G.; Li, Y.S. Expression of a novel OSPGYRP (rice proline-, glycine-and tyrosine-rich protein) gene, which is involved in vesicle trafficking, enhanced cold tolerance in E. coli. Biotechnol. Lett. 2009, 31, 905–910. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Miura, K.; Nagano, K.; Hayanosaito, Y.; Saito, A.; Araki, H.; Kato, A. Chromosomal location of quantitative trait loci for cool tolerance at the booting stage in rice variety Norin-PL8. Breed. Sci. 1995, 45, 337–340. [Google Scholar] [CrossRef]
- Zeng, Y.W.; Zhang, H.L.; Li, Z.C.; Shen, S.Q.; Sun, J.L.; Wang, M.X.; Liao, D.Q.; Liu, X.; Wang, X.K.; Xiao, F.H.; et al. Evaluation of genetic diversity in the rice landraces (Oryza sativa L.) in Yunnan, China. Breed. Sci. 2007, 57, 91–99. [Google Scholar] [CrossRef]
- Zhang, H.L.; Sun, J.L.; Wang, M.X.; Liao, D.Q.; Zeng, Y.W.; Shen, S.Q.; Yu, P.; Mu, P.; Wang, X.K.; Li, Z.C. Genetic structure and phylogeography of rice landraces in Yunnan, China revealed by SSR. Genome 2006, 50, 72–83. [Google Scholar] [CrossRef]
- Dai, L.Y.; Ye, C.R.; Xiong, J.H.; Wang, H.Y. Methods for evaluating cold tolerance in rice. Chin. J. Rice Sci. 1999, 13, 62. [Google Scholar]
- Zeng, Y.W.; Pu, X.Y. Genetic analysis for cold tolerance at booting stage for japonica rice (Oryza sativa L.). Indian J. Genet. 2006, 66, 100–102. [Google Scholar]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from Plant Tissues. In Plant Molecular Biology Manual; Springer: New York, NY, USA, 1989; pp. 73–83. [Google Scholar]
- Ji, Y.T.; Qu, C.Q.; Cao, B.Y. An optimal method of DNA silver staining in polyacrylamide gels. Electrophoresis. 2007, 28, 1173–1175. [Google Scholar] [CrossRef]
- McCouch, S.R.; Temnykh, S.; Lukashova, A.; Coburn, J.; DeClerck, G.; Cartinhour, S.; Harrington, S.; Thomson, M.; Septiningsih, E.; Semon, M.; et al. Microsatellite markers in rice: Abundance, diversity, and applications. In Rice Genetics IV; Khush, G.S., Brar, D.S., Hardy, B., Eds.; Science Publications, New Delhi/IRRI: Los Banos, Philippines, 2001; pp. 117–135. [Google Scholar]
- McCouch, S.R.; Teytelman, L.; Xu, Y.; Lobos, K.B.; Clare, K.; Walton, M.; Fu, B.; Maghirang, R.; Li, Z.; Xing, Y.; et al. Development of 2240 new SSR markers for rice (Oryza sativa L.). DNA Res. 2002, 9, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Temnykh, S.; Declerck, G.; Luashova, A.; Lipovich, L.; Cartinhour, S.; McCouch, S.R. Computational and experimental analysis of microsatellites in rice (Oryza sativa L.): Frequency, length variation, transposon associations, and genetic marker potential. Genome Res. 2001, 11, 1441–1452. [Google Scholar] [CrossRef] [PubMed]
- Temnykh, S.; Park, W.D.; Ayres, N.; Cartinhour, S.; Hauck, N.; Lipovich, L.; Cho, Y.G.; Ishii, T.; McCouch, S.R. Mapping and genome organization of microsatellite sequences in rice (Oryza sativa L.). Theor. Appl. Genet. 2000, 100, 697–712. [Google Scholar] [CrossRef]
- Lincoln, L.; Lander, E. Constructing Genetic Linkage Maps with MAPMAKER/EXP Version 3.0: A Tutorial and Reference Manual; Lander: Cambridge, UK, 1993; pp. 1–9. [Google Scholar]
- Gao, Y.M.; Zhu, J. Mapping QTLs with digenic epistasis under multiple environments and predicting heterosis based on QTL effects. Theor. Appl. Genet. 2007, 115, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J. Mixed model approaches for mapping quantitative trait loci. Hereditas 1998, 20, 137–138. [Google Scholar]
- Wang, D.L.; Zhu, J.; Li, Z.R.; Paterson, A.H. Mapping QTLs with epistatic effects and QTL environment interactions by mixed linear model approaches. Theor. Appl. Genet. 1999, 99, 1255–1264. [Google Scholar] [CrossRef]
- Jagadish, K.S.V.; Craufurd, P.; Shi, W.J.; Oane, R. A phenotypic marker for quantifying heat stress impact during microsporogenesis in rice (Oryza sativa L.). Funct. Plant Biol. 2014, 41, 48–55. [Google Scholar] [CrossRef]
- Mitchell, J.H.; Zulkafli, S.L.; Bosse, J.; Campbell, B.; Snell, P.; Mace, E.S.; Godwin, I.D.; Fukai, S. Rice-cold tolerance across reproductive stages. Crop Pasture Sci. 2016, 67, 823–833. [Google Scholar] [CrossRef]
- Heenan, D.P. Low-temperature induced floret sterility in the rice cultivars Calrose and Inga as influenced by nitrogen supply. Aust. J. Agric. Anim. Husb. 1984, 24, 255–259. [Google Scholar] [CrossRef]
- Suh, J.; Lee, C.; Lee, J.; Kim, J.; Kim, S.; Cho, Y.; Park, S.; Shin, J.; Kim, Y.; Jena, K. Identification of quantitative trait loci for seedling cold tolerance using RILs derived from a cross between japonica and tropical japonica rice cultivars. Euphytica 2012, 184, 101–108. [Google Scholar] [CrossRef]
- Shen, S.Q.; Zeng, Y.W.; Li, S.C.; Wen, G.S.; Pu, X.Y. Mapping cold tolerance gene by near-isogenic lines at booting stage in japonica rice. Chin. J. Rice Sci. 2005, 19, 217–222. [Google Scholar]
- Wang, L.Z.; Wang, L.M.; Xiang, H.T.; Luo, Y.; Li, R.; Li, Z.J.; Wang, C.Y.; Meng, Y. Relationship of photosynthetic efficiency and seed-setting rate in two contrasting rice cultivars under chilling stress. Photosynthetica 2016, 54, 581–588. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Muthurajan, R.; Oane, R.; Wheeler, T.R.; Heuer, S.; Bennett, J.; Craufurd, P.Q. Physiological and proteomic approaches to address heat tolerance during anthesis in rice (Oryza sativa L.). J. Exp. Bot. 2010, 61, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Hayano-Saito, Y.; Maruyama-Funatsuki, W.; Sato, Y.; Kato, A. Physical mapping and putative candidate gene identification of a quantitative trait locus Ctb1 for cold tolerance at the booting stage of rice. Theor. Appl. Genet. 2004, 109, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, K.; Takahashi, M.; Nakazawa, Y. Antibiosis Resistance of QTL Introgressive Soybean Lines to Common Cutworm (Spodoptera litura Fabricius). Crop Sci. 2008, 48, 527–532. [Google Scholar] [CrossRef]
- Suh, J.; Jeung, J.; Lee, J.; Choi, Y.; Yea, J.; Virk, P.; Mackill, D.; Jena, K. Identification and analysis of QTLs controlling cold tolerance at the reproductive stage and validation of effective QTLs in cold-tolerant genotypes of rice (Oryza sativa L.). Theor. Appl. Genet. 2010, 120, 985–995. [Google Scholar] [CrossRef]

 ), BG (
), BG (  ), ILBS (
), ILBS (  ), ULL (
), ULL (  ), RSLL (
), RSLL (  ), FG (
), FG (  ), and 1-2IL (
), and 1-2IL (  ) are located on chromosome number 7.
), BG ( ), ILBS ( ), ULL ( ), RSLL ( ), FG ( ), and 1-2IL ( ) are located on chromosome number 7.
) are located on chromosome number 7.
), BG ( ), ILBS ( ), ULL ( ), RSLL ( ), FG ( ), and 1-2IL ( ) are located on chromosome number 7.

{kind=link}
{kind=link}
| Experimental Locations | ||
|---|---|---|
| Kunming | Aziying | |
| Number of lines | 264 × 2 | 264 × 2 |
| Sowing date | Mid-March | Mid-March |
| Transplant date | Late May | Late May |
| Harvest date | Early October | Early October |
| Average temperature | 16–18 °C | 15–18°C |
| Cold treatment method | Natural low temperature | Cool-water irrigation system |
| Duration of booting to maturity | July to August | July to August |
| Air temperature (booting stage) | 19.0 °C | 17 °C |
| Water temperature | 18.5 ± 0.50 | 17.4 ± 0.30 |
| Morphological Traits | Rate of Seed Setting (RSS) | |||
|---|---|---|---|---|
| K | A | Mean | Difference | |
| AL | 0.19 ** | 0.24 ** | 0.23 ** | 0.02 |
| AW | 0.12 * | 0.17 ** | 0.13 * | 0.07 |
| PHt | 0.06 | 0.14* | 0.09 | 0.04 |
| ET | −0.17 ** | 0.08 | −0.06 | −0.14 * |
| PL | 0.07 | 0.03 | 0.02 | 0.10 |
| FLL | −0.08 | 0.00 | −0.05 | −0.05 |
| FLW | −0.32 ** | 0.29 ** | 0.00 | −0.18 ** |
| ILBS | −0.04 | 0.08 | −0.08 | 0.05 |
| RFLL | −0.11 | −0.17 ** | −0.06 | −0.16 ** |
| RFLW | −0.24 ** | 0.35 ** | 0.02 | −0.04 |
| UIL | 0.27 ** | 0.22 ** | 0.24 ** | 0.20 ** |
| RSLL | −0.23 ** | −0.12 * | −0.17 ** | −0.13 * |
| RSLW | −0.16 ** | 0.31 ** | −0.07 | 0.07 |
| SIL | −0.10 | 0.019 | −0.02 | −0.15 * |
| 1-2IL | 0.23 ** | 0.53 ** | 0.32 ** | 0.16 * |
| PaL | −0.05 | −0.17 ** | −0.07 | −0.03 |
| FG | 0.89 ** | 0.98 ** | 0.89 ** | 0.89 ** |
| BG | −0.91 ** | −0.12 * | −0.66 ** | −0.66 ** |
| NOGP | 0.098 | 0.057 | 0.00 | 0.204 ** |
| Traits | Parents | Near Isogenic Lines (NILs) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Towada | Lijing2 | Standard Deviation | Mean | CV% | Mini/Max | K-S-p | Skewness | Kurtosis | |
| RSS | 0.24 | 0.94 | 0.13 | 0.54 ± 0.01 | 24.29 | 0.06/0.92 | 0.53 | −0.3 | 0.24 |
| FG | 24.6 | 123.7 | 23.16 | 83.49 ± 1.43 | 27.74 | 7.60/152.50 | 0.88 | 0.02 | 0.33 |
| BG | 80.3 | 7.5 | 20.42 | 69.63 ± 1.26 | 29.33 | 11.40/133.40 | 0.66 | 0.36 | −0.07 |
| ULL | 21.14 | 25.5 | 1.39 | 21.99 ± 0.09 | 6.32 | 18.04/25.88 | 0.83 | 0.23 | 0.07 |
| 1-2IL | 2.38 | 4.09 | 0.36 | 2.36 ± 0.02 | 15.19 | 1.33/3.45 | 0.69 | −0.15 | −0.12 |
| RSLL | 30.8 | 35.18 | 2.69 | 36.93 ± 0.17 | 7.28 | 30.87/47.58 | 0.18 | 0.44 | 0.34 |
| McCouch Locus ID | Motif | Number of Repeats | Forward Primer | Reverse Primer |
|---|---|---|---|---|
| RM21344 | AT | 12 | GGATGTTGTTCTAACCCGTCAGG | CGAACTCAACAGACTACCCATACCC |
| RM21345 | AT | 15 | GCATGCTAAGCTGTAGAAGTTAGTGG | GCTACATGTCACCGATCAGACC |
| RM21351 | AT | 15 | GGTGGTGTTTGTAAGTGTTTCACG | CGAACATGAACATAGGTCGTTGC |
| RM21353 | AT | 10 | GGAAACCACATGCTTGATGC | CACTCCTTATATGGATGGTTAGGG |
| RM21361 | AGAT | 5 | GATGGGAAGAGACGAGAGTTGACC | TAGGAGTGATACGTGGCGTCTTAGG |
| RM8036 | AT | 37 | ATGGTTTGGAGCTTGAACTGTCC | GGTTAGGAGCAGTGGGAATTTGG |
| RM21476 | AGG | 10 | GACGCCGACGATCTCCATCTCC | GCAAGAGTACTATGCGGCGGAAGC |
| RM21478 | AAG | 28 | TAACACAGTTCTTCTCGCAACG | AAGTTCCCTTGTGTGATTGACC |
| RM3755 | AG | 17 | TGTGGACAACCTCAACTGAAAGC | CATAATCACCAACATCGGAGAAGC |
| RM5499 | AG | 25 | GGACGAAAGGGTATTTGATTGG | CCTCAAGGTGGTCTCCTTCTCC |
| Traits | Chromosome Number | Interval Distance | Site (cM) a | LOD | Additive Effect (%2) b | H2(A) % c |
|---|---|---|---|---|---|---|
| RSS | 7 | RM21353–RM21345 | 0.06 | 3.94 | −0.04 | 8.36 |
| RSS | 7 | RM21476–RM21478 | 0.00 | 3.03 | −0.03 | 5.7 |
| 1-2IL | 7 | RM21345–RM21344 | 0.06 | 2.60 | −0.11 | 7.31 |
| BG | 7 | RM21353–RM21345 | 0.06 | 3.60 | 5.98 | 7.67 |
| BG | 7 | RM21476–RM21478 | 0.00 | 2.06 | 4.22 | 3.92 |
| RSLL | 7 | RM21345–RM21344 | 0.48 | 2.83 | 0.65 | 5.79 |
| FG | 7 | RM21353–RM21345 | 0.04 | 2.94 | −6.22 | 6.63 |
| FG | 7 | RM3755–RM8036 | 0.00 | 2.71 | −5.45 | 5.2 |
| ILBS | 7 | RM3755–RM8036 | 0.08 | 2.03 | −0.5 | 4.37 |
| ULL | 7 | RM5499–RM21353 | 0.00 | 3.14 | −0.34 | 5.71 |
| ULL | 7 | RM21476–RM21478 | 0.00 | 3.67 | −0.38 | 6.87 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Du, J.; Pu, X.; Kazim Ali, M.; Yang, X.; Duan, C.; Ren, M.; Li, X.; Zeng, Y. Near-Isogenic Lines of Japonica Rice Revealed New QTLs for Cold Tolerance at Booting Stage. Agronomy 2019, 9, 40. https://doi.org/10.3390/agronomy9010040
Sun Z, Du J, Pu X, Kazim Ali M, Yang X, Duan C, Ren M, Li X, Zeng Y. Near-Isogenic Lines of Japonica Rice Revealed New QTLs for Cold Tolerance at Booting Stage. Agronomy. 2019; 9(1):40. https://doi.org/10.3390/agronomy9010040
Chicago/Turabian StyleSun, Zhenghai, Juan Du, Xiaoying Pu, Muhammad Kazim Ali, Xiaomeng Yang, Chengli Duan, Meirong Ren, Xia Li, and Yawen Zeng. 2019. "Near-Isogenic Lines of Japonica Rice Revealed New QTLs for Cold Tolerance at Booting Stage" Agronomy 9, no. 1: 40. https://doi.org/10.3390/agronomy9010040