Delivery of Inoculum of Rhizophagus irregularis via Seed Coating in Combination with Pseudomonas libanensis for Cowpea Production

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Plant
2.2. Bacterial Strain
2.3. Arbuscular Mycorrhizal Fungal Inoculum and Seed Coating
2.4. Microcosm Experiments
2.4.1. Plant Biomass
2.4.2. Pigment Estimation
2.4.3. Mycorrhizal Colonization
2.4.4. Bacterial Analysis
2.5. Statistical Analysis
3. Results
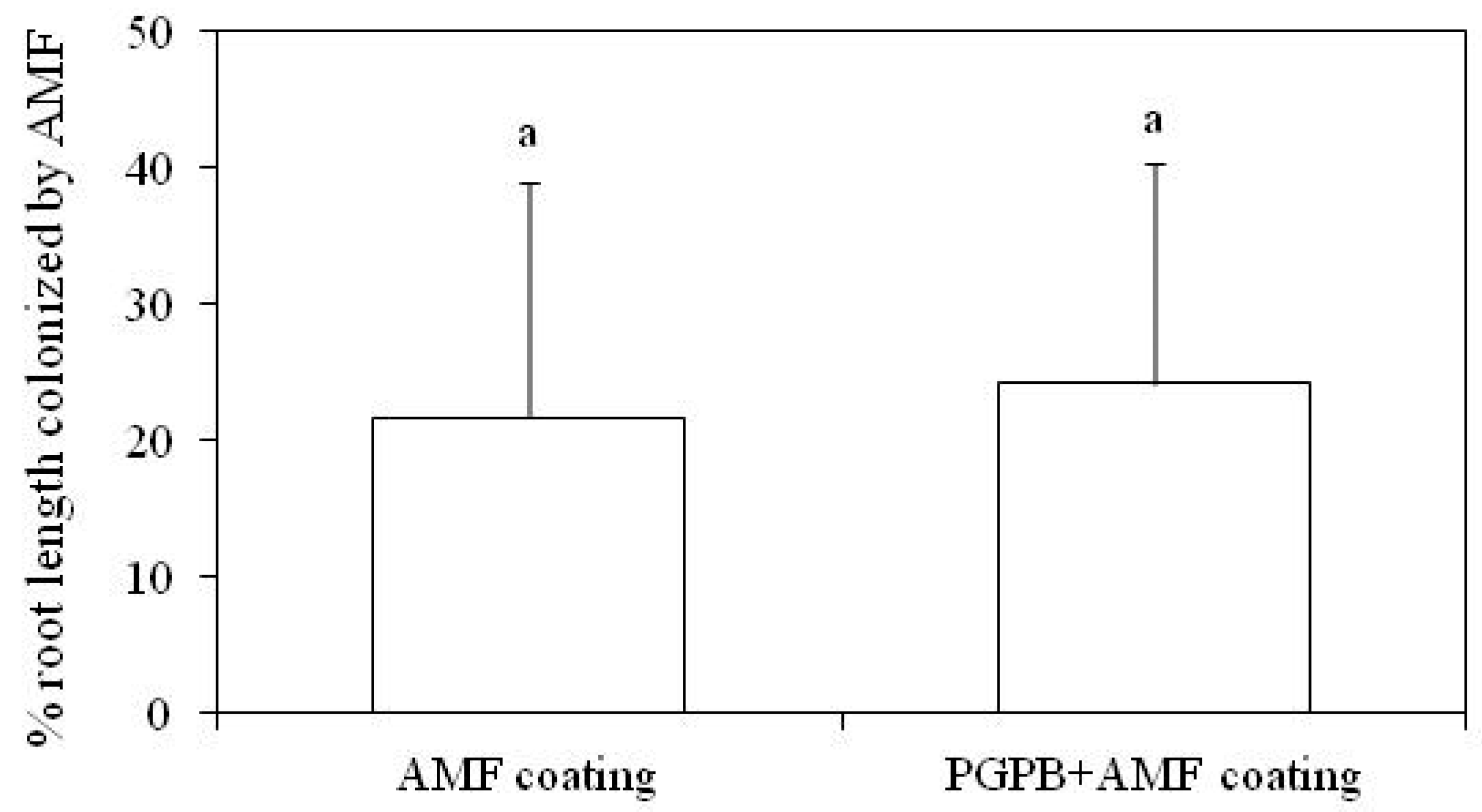
3.1. AMF Colonization
3.2. Bacterial Colonization
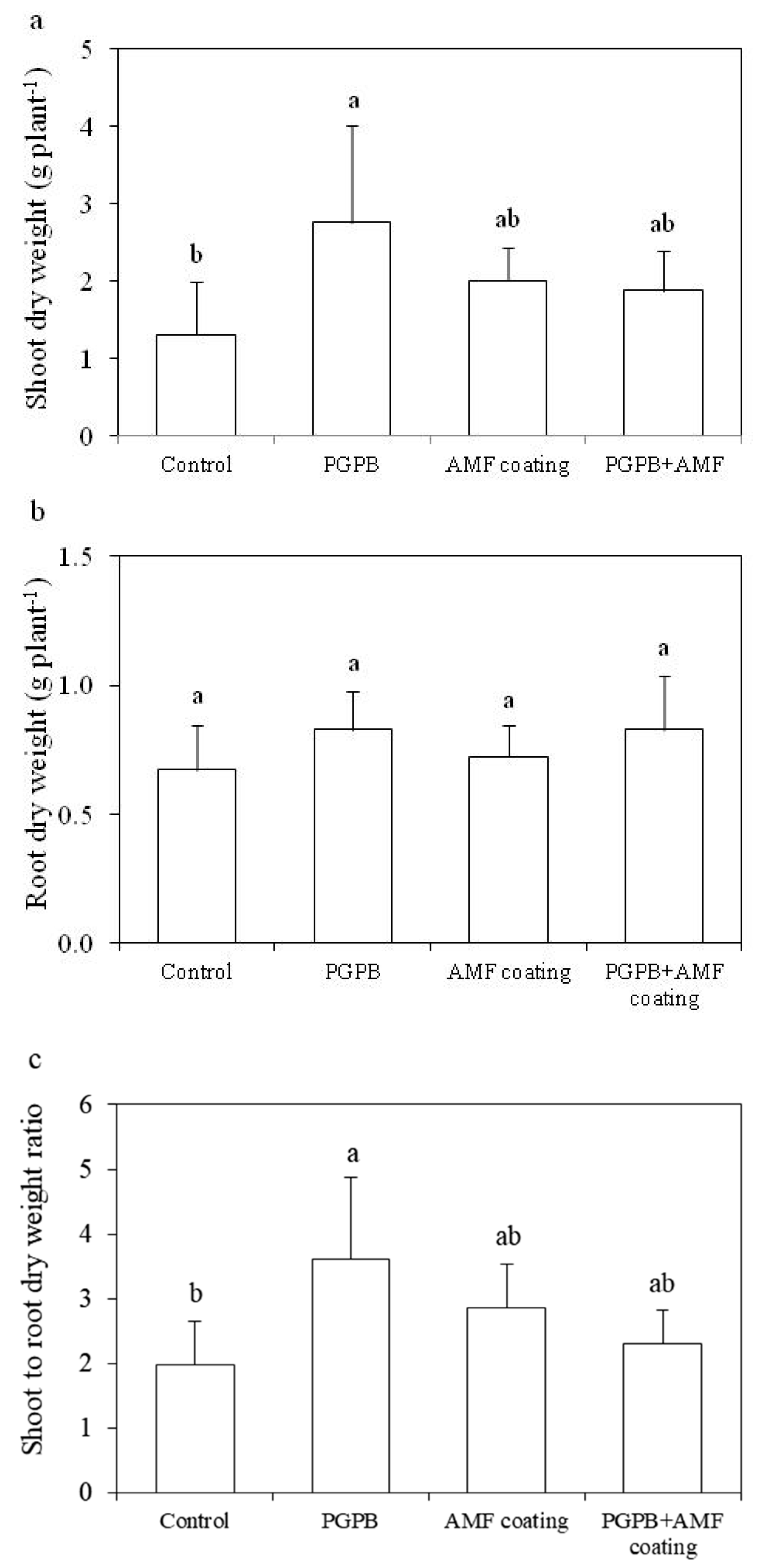
3.3. Plant Biomass and Chlorophyll Contents
3.4. Effects of Microbes on Seed Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fitter, A.H.; Helgason, T.; Hodge, A. Nutritional exchanges in the arbuscular mycorrhizal symbiosis: Implications for sustainable agriculture. Fungal Biol. Rev. 2011, 25, 68–72. [Google Scholar] [CrossRef]
- Ma, Y.; Prasad, M.N.V.; Rajkumar, M.; Freitas, H. Plant growth promoting rhizobacteria and endophytes accelerate phytoremediation of metalliferous soils. Biotechnol. Adv. 2011, 29, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Oliveira, R.S.; Freitas, H.; Zhang, C. Biochemical and molecular mechanisms of plant-microbe-metal interactions: Relevance for phytoremediation. Front. Plant Sci. 2016, 7, 918. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.S.; Ma, Y.; Rocha, I.; Carvalho, M.F.; Vosátka, M.; Freitas, H. Arbuscular mycorrhizal fungi are an alternative to the application of chemical fertilizer in the production of the medicinal and aromatic plant Coriandrum sativum L. J. Toxicol. Environ. Health A 2016, 79, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.S.; Rocha, I.; Ma, Y.; Vosátka, M.; Freitas, H. Seed coating with arbuscular mycorrhizal fungi as an ecotechnological approach for sustainable agricultural production of common wheat (Triticum aestivum L.). J. Toxicol. Environ. Health A 2016, 79, 329–337. [Google Scholar] [CrossRef]
- Gonçalves, A.; Goufo, P.; Barros, A.; Domínguez-Perles, R.; Trindade, H.; Rosa, E.A.; Ferreira, L.; Rodrigues, M. Cowpea (Vigna unguiculata L. Walp), a renewed multipurpose crop for a more sustainable agri-food system: Nutritional advantages and constraints. J. Sci. Food Agric. 2016, 96, 2941–2951. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H.M. Drought stress in plants: An overview. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer Press: Berlin/Heidelberg, Germany, 2012; pp. 1–33. [Google Scholar]
- Kareem, K.T.; Taiwo, M.A. Interactions of viruses in cowpea: Effects on growth and yield parameters. Virol. J. 2007, 4, 1–15. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.; Streitwolf-Engel, R.; Riedl, R.; Siegrist, S.; Neudecker, A.; Ineichen, K.; Boller, T.; Wiemken, A.; Sanders, I.R. The mycorrhizal contribution to plant productivity, plant nutrition and soil structure in experimental grassland. New Phytol. 2006, 172, 739–752. [Google Scholar] [CrossRef] [Green Version]
- Reinhold-Hurek, B.; Hurek, T. Living inside plants: Bacterial endophytes. Curr. Opin. Plant Biol. 2011, 14, 435–443. [Google Scholar] [CrossRef]
- Ehsanfar, S.; Mdarre, S. Crop protection by seed coating. Commun. Agric. Appl. Biol. Sci. 2005, 70, 225–229. [Google Scholar]
- Pedrini, S.; Merritt, D.J.; Stevens, J.; Dixon, K. Seed coating: Science or marketing spin? Trends Plant Sci. 2017, 22, 106–116. [Google Scholar] [CrossRef]
- Ma, Y.; Rajkumar, M.; Zhang, C.; Freitas, H. Inoculation of Brassica oxyrrhina with plant growth promoting bacteria for the improvement of heavy metal phytoremediation under drought conditions. J. Hazard. Mater. 2016, 320, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Porter, W. The “most probable number” method for enumerating infective propagules of vesicular arbuscular mycorrhizal fungi in soil. Aust. J. Soil Res. 1979, 17, 515–519. [Google Scholar] [CrossRef]
- Scott, J.M.; Hill, C.B.; Jessop, R.S. Growth chamber study of phosphorus applied as drilled granules or as seed coatings to wheat sown in soils differing in P-sorption capacity. Fertil. Res. 1991, 29, 281–287. [Google Scholar] [CrossRef]
- Moran, R. Formulae for determination of chlorophyllous pigments extracted with N,N-Dimethylformamide. Plant Physiol. 1982, 69, 1376–1381. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.; Wellburn, A. Determination of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 603, 591–592. [Google Scholar] [CrossRef]
- Koske, R.E.; Gemma, J.N. A modified procedure for staining roots to detect VA mycorrhizas. Mycol. Res. 1989, 92, 486–505. [Google Scholar] [CrossRef]
- Mcgonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 1990, 155, 495–501. [Google Scholar] [CrossRef]
- Widmer, F.; Seidler, R.J.; Gillevet, P.M.; Watrud, L.S.; Di Giovanni, G.D. A highly selective PCR protocol for detecting 16S rRNA genes of the genus Pseudomonas (sensu stricto) in environmental samples. Appl. Environ. Microbiol. 1998, 64, 2545–2553. [Google Scholar]
- Rajwar, A.; Sahgal, M. Phylogenetic relationships of Fluorescent pseudomonads deduced from the sequence analysis of 16S rRNA, Pseudomonas-specific and rpoD genes. 3 Biotech 2016, 6, 80. [Google Scholar] [CrossRef]
- Sinha, S.K. Yield, yield components and plant ideotype in food legumes. In: Food legume crops: Improvement and production. FAO Plant Prod. Protect. Ser. 1977, 9, 123–131. [Google Scholar]
- Yang, D.; Wang, N.; Yan, X.; Shi, J.; Zhang, M.; Wang, Z.; Yuan, H. Microencapsulation of seed-coating tebuconazole and its effects on physiology and biochemistry of maize seedlings. Colloids Surf. B Biointerfaces 2014, 114, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Omirou, M.; Fasoula, D.A.; Ioannides, I.M. Bradyrhizobium inoculation alters indigenous AMF community assemblages and interacts positively with AMF inoculum to improve cowpea performance. Appl. Soil Ecol. 2016, 108, 381–389. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.; Kim, K.; Subramanian, P.; Senthilkumar, M.; Anandham, R.; Sa, T. Arbuscular mycorrhizal fungi and associated bacteria isolated from salt-affected soil enhances the tolerance of maize to salinity in coastal reclamation soil. Agric. Ecosyst. Environ. 2016, 231, 233–239. [Google Scholar] [CrossRef]
- Crecchio, C.; Curci, M.; Mininni, R.; Ricciuti, P.; Ruggiero, P. Short-term effects of municipal solid waste compost amendments on soil carbon and nitrogen content, some enzyme activities and genetic diversity. Biol. Fertil. Soils 2001, 34, 311–318. [Google Scholar] [CrossRef]
- Johansson, J.; Paul, L.R.; Finlay, R.D. Microbial interactions in the mycorrhizosphere and their significance for sustainable agriculture. FEMS Microbiol. Ecol. 2004, 48, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gryndler, M.; Hrselová, H.; Sudová, R.; Gryndlerová, H.; Rezácová, V.; Merhautová, V. Hyphal growth and mycorrhiza formation by the arbuscular fungus Glomus calaroideum BEG 23 is stimulated by humic substances. Mycorrhiza 2005, 15, 483–488. [Google Scholar] [CrossRef]
- Genre, A.; Russo, G. Does a common pathway transduce symbiotic signals in plant-microbe interactions? Front. Plant Sci. 2016, 7, 96. [Google Scholar] [CrossRef]
- Bolandnazar, S.; Aliasgarzad, N.; Neishabury, M.R.; Chaparzadeh, N. Mycorrhizal colonization improves onion (Allium cepa L.) yield and water use efficiency under water deficit condition. Sci. Hortic. 2007, 114, 11–15. [Google Scholar] [CrossRef]
- Pellegrino, E.; Bedini, S. Enhancing ecosystem services in sustainable agriculture: Biofertilization and biofortification of chickpea (Cicer arietinum L.) by arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2014, 68, 429–439. [Google Scholar] [CrossRef]
- Allah, E.F.A.; Hashem, A.; Alqarawi, A.A.; Bahkali, A.H.; Alwhibi, M.S. Enhancing growth performance and systemic acquired resistance of medicinal plant Sesbania sesban (L.) Merr using arbuscular mycorrhizal fungi under salt stress. Saudi J. Biol. Sci. 2015, 22, 274–283. [Google Scholar]
- Li, J.F.; He, X.H.; Li, H.; Zheng, W.J.; Liu, J.F.; Wang, M.Y. Arbuscular mycorrhizal fungi increase growth and phenolics synthesis in Poncirus trifoliate under iron deficiency. Sci. Hortic. 2015, 183, 87–92. [Google Scholar] [CrossRef]
- Hristozkova, M.; Geneva, M.; Stancheva, I.; Boychinova, M.; Djonova, E. Contribution of arbuscular mycorrhizal fungi in attenuation of heavy metal impact on Calendula officinalis development. Appl. Soil Ecol. 2016, 101, 57–63. [Google Scholar] [CrossRef]
- Oliveira, R.S.; Carvalho, P.; Marques, G.; Ferreira, L.; Nunes, M.; Rocha, I.; Ma, Y.; Carvalho, M.F.; Vosátka, M.; Freitas, H. Increased protein content of chickpea (Cicer arietinum L.) inoculated with arbuscular mycorrhizal fungi and nitrogen-fixing bacteria under water deficit conditions. J. Sci. Food Agric. 2017, 97, 4379–4385. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.S.; Carvalho, P.; Marques, G.; Ferreira, L.; Pereira, S.; Nunes, M.; Rocha, I.; Ma, Y.; Carvalho, M.F.; Vosátka, M.; et al. Improved grain yield of cowpea (Vigna unguiculata) under water deficit after inoculation with Bradyrhizobium elkanii and Rhizophagus irregularis. Crop Pasture Sci. 2017, 68, 1052–1059. [Google Scholar] [CrossRef]
- Muthukumar, T.; Udaiyan, K. Arbuscular mycorrhizas in cycads of southern India. Mycorrhiza 2002, 12, 213–217. [Google Scholar] [PubMed]
- Johnson, N.C.; Graham, J.H.; Smith, F.A. Functioning of mycorrhizal associations along the mutualism–parasitism continuum. New Phytol. 1997, 135, 575–585. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by Fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–331. [Google Scholar] [CrossRef]
- Rosas, S.B.; Avanzini, G.; Carlier, E.; Pasluosta, C.; Pastor, N.; Rovera, M. Root colonization and growth promotion of wheat and maize by Pseudomonas aurantiaca SR1. Soil Biol. Biochem. 2009, 41, 1802–1806. [Google Scholar] [CrossRef]
- Barrow, J.R.; Lucero, M.E.; Reyes-Vera, I.; Havstad, K.M. Do symbiotic microbes have a role in plant evolution, performance and response to stress? Commun. Integr. Biol. 2008, 1, 69–73. [Google Scholar] [CrossRef]
- Bashan, Y.; Dubrovsky, J.G. Azospirillum spp. participation in dry matter partitioning in grasses at the whole plant level. Biol. Fertil. Soils 1996, 23, 435–440. [Google Scholar] [CrossRef]
- Nouri, E.; Breuillin-Sessoms, F.; Feller, U.; Reinhardt, D. Correction: Phosphorus and nitrogen regulate arbuscular mycorrhizal symbiosis in Petunia hybrida. PLoS ONE 2015, 10, e0127472. [Google Scholar] [CrossRef] [PubMed]
- Abayomi, Y.A.; Ajibade, T.V.; Sammuel, O.F.; Saadudeen, B.F. Growth and yield responses of cowpea (Vigna unguiculata (L.) Walp) genotypes to nitrogen fertilizer (NPK) application in the Southern Guinea Savanna zone of Nigeria. Asian J. Plant Sci. 2008, 7, 170–176. [Google Scholar] [CrossRef]
- Valverde, A.; Burgos, A.; Fiscella, T.; Rivas, R.; Velázquez, E.; Rodríguez-Barrueco, C.; Cervantes, E.; Chamber, M.; Igual, J.M. Differential effects of coinoculations with Pseudomonas jessenii PS06 (a phosphate-solubilizing bacterium) and Mesorhizobium ciceri C-2/2 strains on the growth and seed yield of chickpea under greenhouse and field conditions. Plant Soil 2006, 287, 43–50. [Google Scholar] [CrossRef]
- Parra-Cota, F.I.; Peña-Cabriales, J.J.; De Los Santos-villalobos, S.; Martínez-Gallardo, N.A.; Délano-Frier, J.P. Burkholderia ambifaria and B. caribensis promote growth and increase yield in grain amaranth (Amaranthus cruentus and A. hypochondriacus) by improving plant nitrogen uptake. PLoS ONE 2014, 9, e88094. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Characteristic | Parameter | Unit | P. libanensis TR1 |
|---|---|---|---|
| Abiotic stress resistance | Salt tolerance | % | 8 |
| Osmotic tolerance * | nq | + | |
| Temperature tolerance | °C | 4–38 | |
| Plant growth promoting traits | ACC deaminase production | μm α-KB mg−1 h−1 protein | 34.2 ± 6.7 |
| P solubilization | nq | + | |
| IAA production | mg L−1 | 88.2 ± 5.6 | |
| Siderophore | CAS: mm | 1.0 ± 0.1 | |
| N2 fixation | nq | + | |
| NH3 production | nq | + |
| Treatment | Chlorophyll a (mg g −1) | Chlorophyll b (mg g −1) | Chlorophyll a + b (mg g −1) | Chlorophyll a/b Ratio |
|---|---|---|---|---|
| Control | 1.89 ± 0.43 a | 3.04 ± 0.70 a | 4.99 ± 1.12 a | 0.63 ± 0.06 a |
| PGPB | 2.24 ± 0.22 a | 3.50 ± 0.41 a | 5.80 ± 0.64 a | 0.64 ± 0.02 a |
| AMF coating | 2.21 ± 0.43 a | 3.38 ± 0.54 a | 5.65 ± 0.95 a | 0.65 ± 0.04 a |
| PGPB + AMF coating | 2.33 ± 0.38 a | 3.53 ± 0.49 a | 5.93 ± 0.88 a | 0.66 ± 0.03 a |
| Treatment | Pod Number | Seed Number | Seed Weight | * Seed Yield |
|---|---|---|---|---|
| Per Plant | Per Pod | Per Seed (g) | Per Plant (g) | |
| Control | 2.3 ± 0.5 a | 3.6 ± 0.7 a | 0.18 ± 0.0 a | 1.43 ± 0.3 b |
| PGPB | 2.6 ± 1.1 a | 3.5 ± 0.9 a | 0.22 ± 0.0 a | 2.18 ± 0.4 a |
| AMF coating | 2.3 ± 0.5 a | 2.5 ± 1.5 a | 0.18 ± 0.1 a | 1.23 ± 0.3 b |
| PGPB + AMF coating | 2.4 ± 0.9 a | 3.7 ± 1.8 a | 0.17 ± 0.1 a | 1.19 ± 0.5 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Látr, A.; Rocha, I.; Freitas, H.; Vosátka, M.; Oliveira, R.S. Delivery of Inoculum of Rhizophagus irregularis via Seed Coating in Combination with Pseudomonas libanensis for Cowpea Production. Agronomy 2019, 9, 33. https://doi.org/10.3390/agronomy9010033
Ma Y, Látr A, Rocha I, Freitas H, Vosátka M, Oliveira RS. Delivery of Inoculum of Rhizophagus irregularis via Seed Coating in Combination with Pseudomonas libanensis for Cowpea Production. Agronomy. 2019; 9(1):33. https://doi.org/10.3390/agronomy9010033
Chicago/Turabian StyleMa, Ying, Aleš Látr, Inês Rocha, Helena Freitas, Miroslav Vosátka, and Rui S. Oliveira. 2019. "Delivery of Inoculum of Rhizophagus irregularis via Seed Coating in Combination with Pseudomonas libanensis for Cowpea Production" Agronomy 9, no. 1: 33. https://doi.org/10.3390/agronomy9010033