Stem Cells Derived from Lipoma and Adipose Tissue—Similar Mesenchymal Phenotype but Different Differentiation Capacity Governed by Distinct Molecular Signature


Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Isolation and Cultivation of Mesenchymal Stem Cells
2.3. Differentiation of Cells
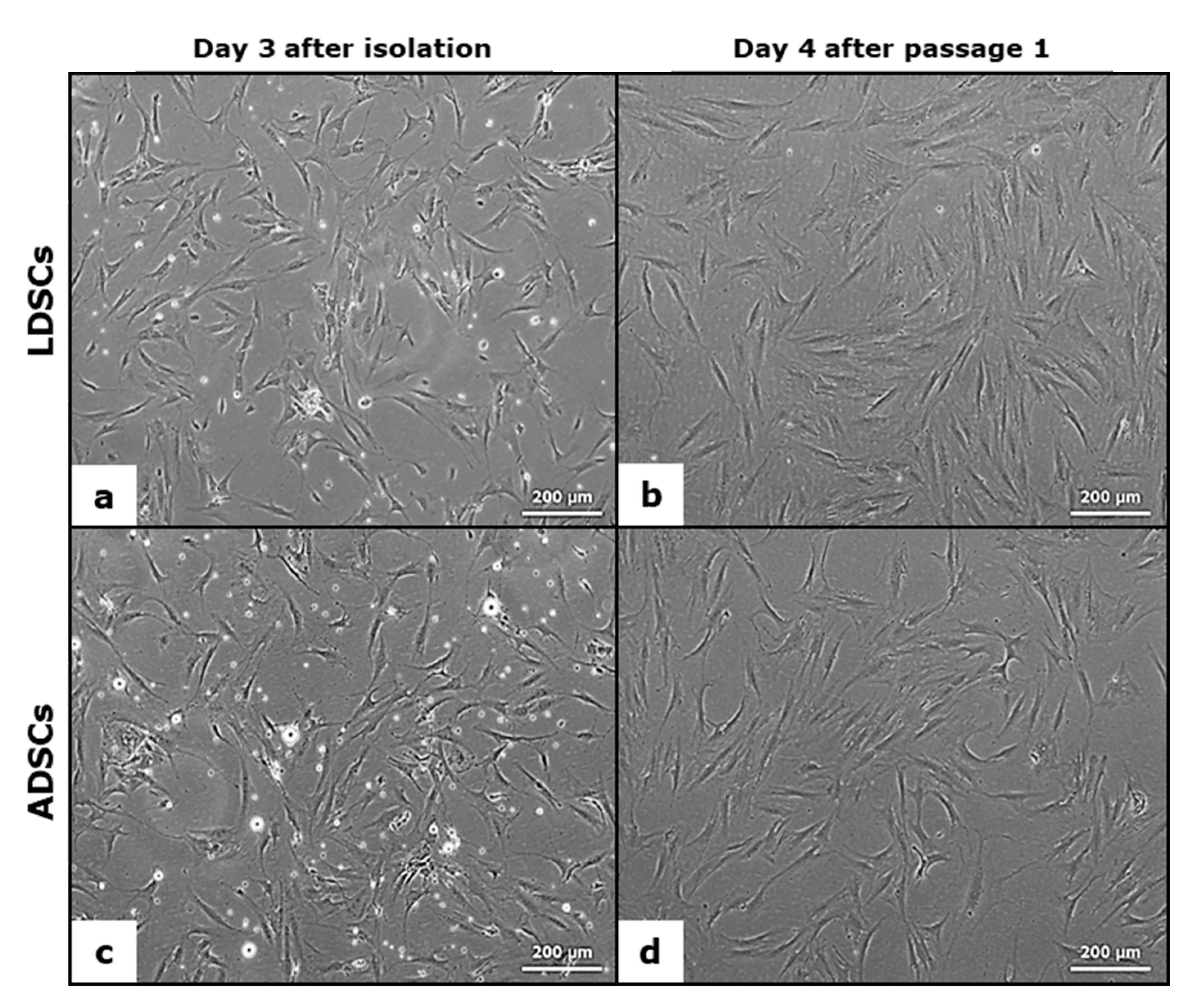
2.4. Light Microscopy
2.5. Flow Cytometry
2.6. RNA Isolation and Reverse Transcription
2.7. Real Time PCR
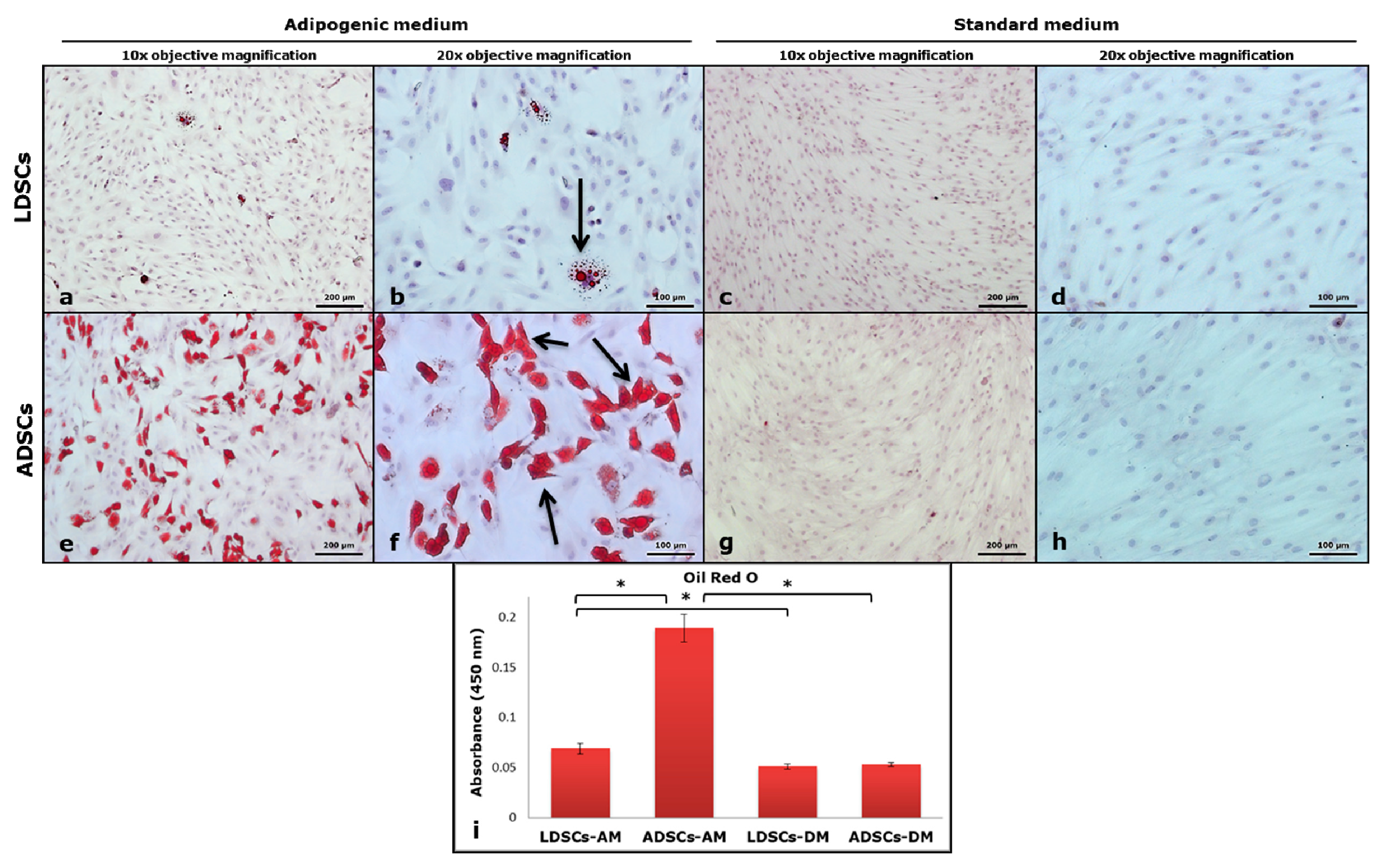
2.8. Oil Red O Staining
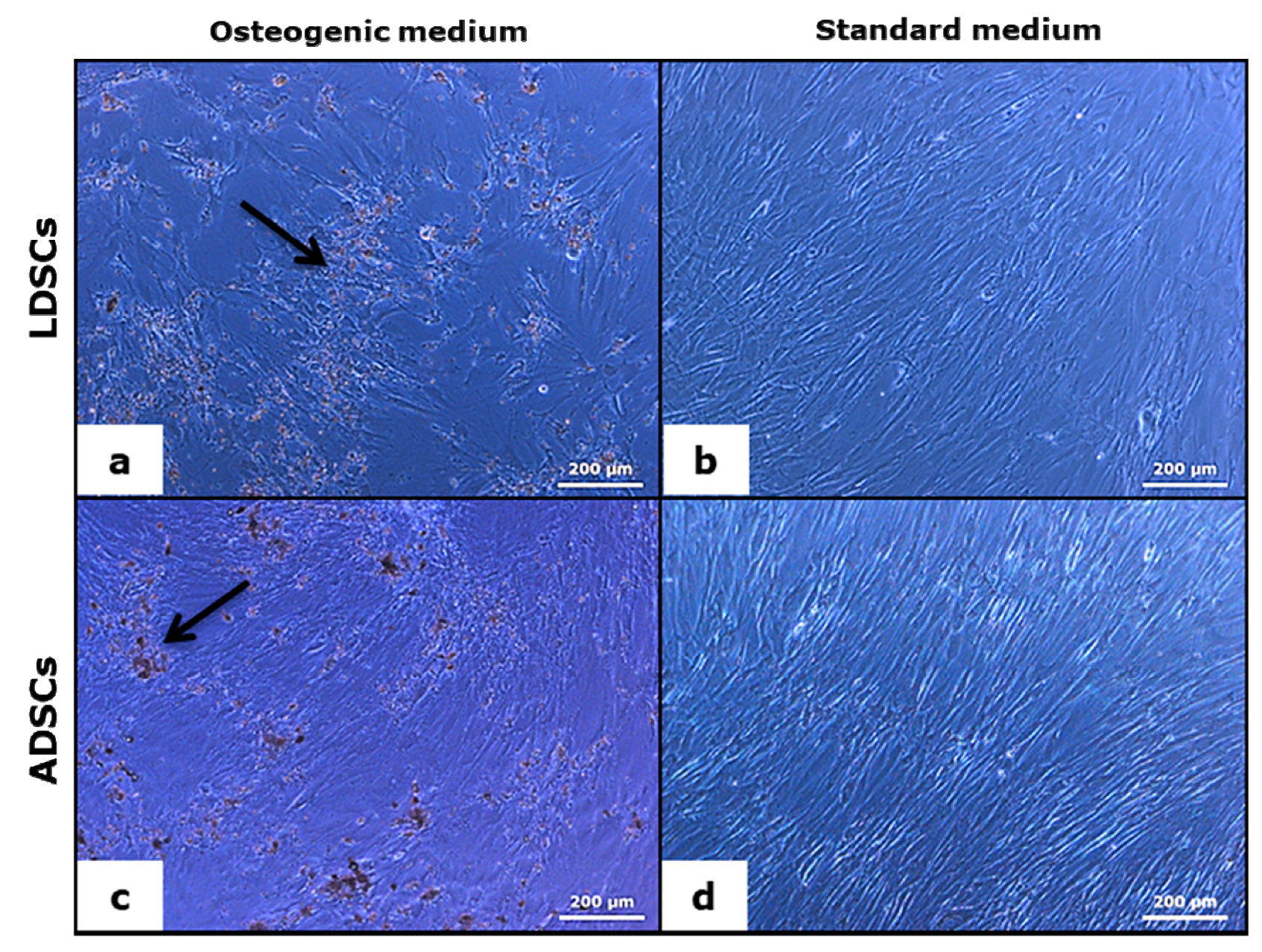
2.9. Cytochemical Staining for Calcium Deposits
2.10. Immunocytochemistry
2.11. ELISA Assays
2.12. Statistical Analysis
3. Results
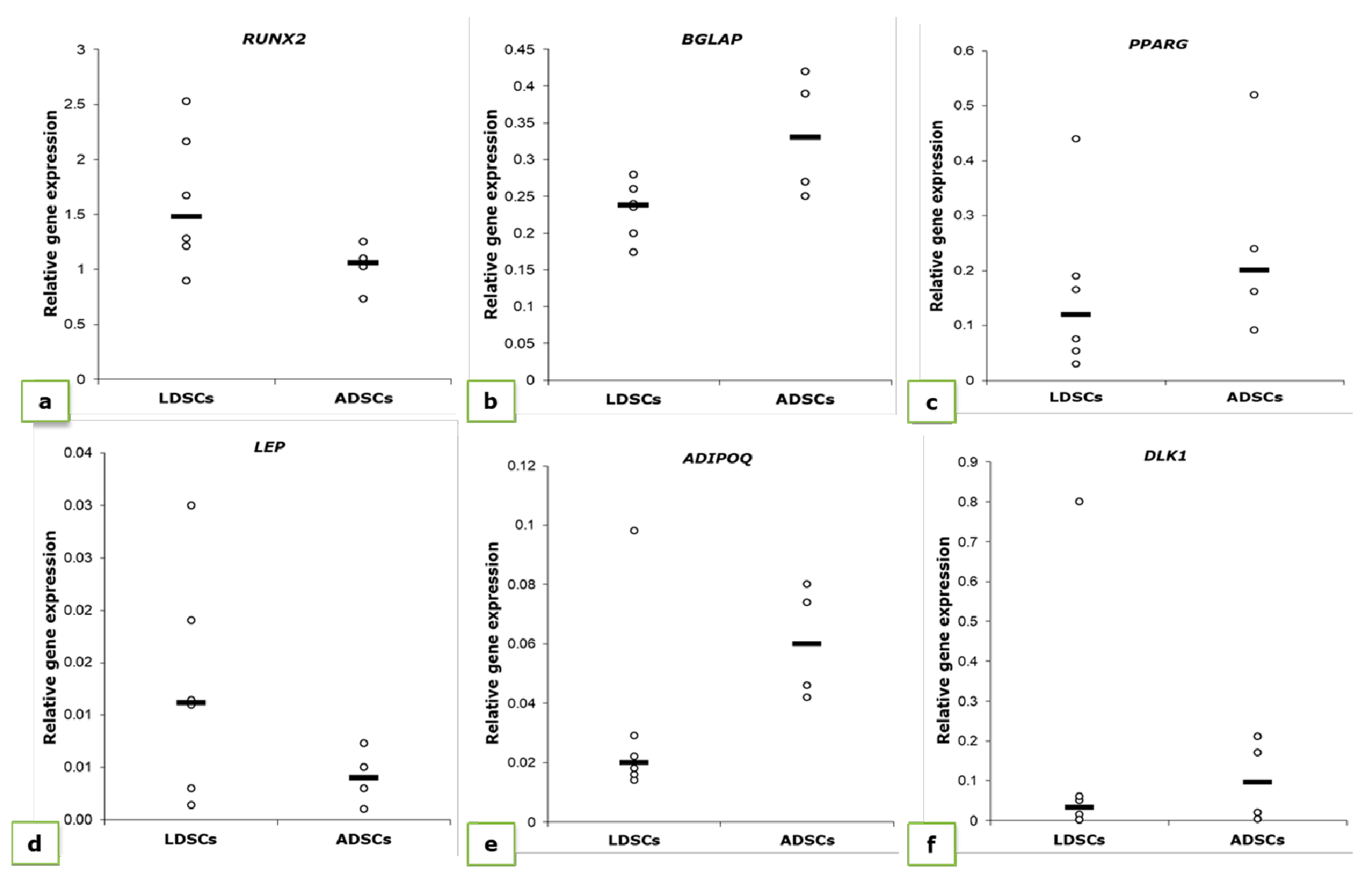
3.1. Analysis of Mesenchymal Stem Cell Phenotype
3.2. Adipogenic Differentiation
3.3. Osteogenic Differentiation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mizuno, H.; Tobita, M.; Uysal, A.C. Concise review: Adipose-derived stem cells as a novel tool for future regenerative medicine. Stem Cells 2012, 30, 804–810. [Google Scholar] [CrossRef]
- Bunnell, B.A.; Flaat, M.; Gagliardi, C.; Patel, B.; Ripoll, C. Adipose-derived stem cells: Isolation, expansion and differentiation. Methods 2008, 45, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Gimble, J.M.; Katz, A.J.; Bunnell, B.A. Adipose-derived stem cells for regenerative medicine. Circ. Res. 2007, 100, 1249–1260. [Google Scholar] [CrossRef]
- Baer, P.C.; Geiger, H. Adipose-derived mesenchymal stromal/ stem cells: Tissue localization, characterization, and heterogeneity. Stem Cells Int. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Cvetković, V.J.; Najdanović, J.G.; Vukelić-Nikolić, M.Đ.; Stojanović, S.; Najman, S.J. Osteogenic potential of in vitro osteo-induced adipose-derived mesenchymal stem cells combined with platelet-rich plasma in an ectopic model. Int. Orthop. 2015, 39, 2173–2180. [Google Scholar] [CrossRef] [PubMed]
- Najdanović, J.G.; Cvetković, V.J.; Stojanović, S.; Vukelić-Nikolić, M.Đ.; Stanisavljević, M.N.; Živković, J.M.; Najman, S.J. The influence of adipose-derived stem cells induced into endothelial cells on ectopic vasculogenesis and osteogenesis. Cell. Mol. Bioeng. 2015, 8, 577–590. [Google Scholar] [CrossRef]
- Najman, S.J.; Cvetković, V.J.; Najdanović, J.G.; Stojanović, S.; Vukelić-Nikolić, M.Đ.; Vučković, I.; Petrović, D. Ectopic osteogenic capacity of freshly isolated adipose-derived stromal vascular fraction cells supported with platelet-rich plasma: A simulation of intraoperative procedure. J. Cranio Maxillofac. Surg. 2016, 44, 1750–1760. [Google Scholar] [CrossRef] [PubMed]
- Omonte, S.V.; de Andrade, B.A.; Leal, R.M.; Capistrano, H.M.; Souza, P.E.; Horta, M.C. Osteolipoma: A rare tumor in the oral cavity. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2016, 122, e8–e13. [Google Scholar] [CrossRef]
- Mohammed, U.; Samaila, M.O.; Abubakar, M. Pattern of adipose tissue tumors in Ahmadu Bello University Teaching Hospital, Zaria, Nigeria. Ann. Niger. Med. 2014, 8, 8–10. [Google Scholar] [CrossRef]
- Mentzel, T.; Fletcher, C.D. Lipomatous tumours of soft tissues: An update. Virchows Arch. 1995, 427, 353–363. [Google Scholar] [CrossRef]
- Marques, M.C.; Garcia, H. Lipomatous Tumors. In Imaging of Soft Tissue Tumors; De Schepper, A.M., Parizel, P.M., Ramon, F., De Beuckeleer, L., Vandevenne, J.E., Eds.; Springer: Berlin Heidelberg, Germany, 1997; pp. 191–207. ISBN 978-3-662-07859-4. [Google Scholar]
- Ingari, J.V.; Faillace, J.J. Benign tumors of fibrous tissue and adipose tissue in the hand. Hand Clin. 2004, 20, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.M.; Chang, H.W.; Wang, K.H.; Kao, A.P.; Chang, C.C.; Wen, C.H.; Lai, C.S.; Lin, S.D. Isolation and identification of mesenchymal stem cells from human lipoma tissue. Biochem. Biophys. Res. Commun. 2007, 361, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Suga, H.; Eto, H.; Inoue, K.; Aoi, N.; Kato, H.; Araki, J.; Higashino, T.; Yoshimura, K. Cellular and molecular features of lipoma tissue: Comparison with normal adipose tissue. Br. J. Dermatol. 2009, 161, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Tremp, M.; Menzi, N.; Tchang, L.; di Summa, P.G.; Schaefer, D.J.; Kalbermatten, D.F. Adipose-derived stromal cells from lipomas: Isolation, characterisation and review of the literature. Pathobiology 2016, 83, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Makiguchi, T.; Terashi, H.; Hashikawa, K.; Yokoo, S.; Kusaka, J. Osteolipoma in the glabella: Pathogenesis associated with mesenchymal lipoma-derived stem cells. J. Craniofac. Surg. 2013, 24, 1310–1313. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.W.; Gao, J.H.; Lu, F.; Zheng, X.D. The differences between adipose tissue derived stem cells and lipoma mesenchymal stem cells incharacteristics. Zhonghua Zheng Xing Wai Ke Za Zhi 2010, 26, 125–132. (In Chinese) [Google Scholar] [PubMed]
- Val-Bernal, J.F.; Val, D.; Garijo, M.F.; Vega, A.; González-Vela, M.C. Subcutaneous ossifying lipoma: Case report and review of the literature. J. Cutan. Pathol. 2007, 34, 788–792. [Google Scholar] [CrossRef]
- Sunohara, M.; Ozawa, T.; Morimoto, K.; Tateishi, C.; Ishii, M. Lipoma with bone and cartilage components in the left axilla of a middle-aged woman. Aesthet. Plast. Surg. 2012, 36, 1164–1167. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Weissgerber, T.L.; Milic, N.M.; Winham, S.J.; Garovic, V.D. Beyond bar and line graphs: Time for a new data presentation paradigm. PLoS Biol. 2015, 13, e1002128. [Google Scholar] [CrossRef]
- Schipper, B.M.; Marra, K.G.; Zhang, W.; Donnenberg, A.D.; Rubin, J.P. Regional anatomic and age effects on cell function of human adipose-derived stem cells. Ann. Plast. Surg. 2008, 60, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Kleineidam, B.; Kleinheinz, J. Regenerative potential of human adipose-derived stromal cells of various origins. J. Cranio Maxillofac. Surg. 2015, 43, 2144–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhery, M.S.; Badowski, M.; Muise, A.; Pierce, J.; Harris, D.T. Donor age negatively impacts adipose tissue-derived mesenchymal stem cell expansion and differentiation. J. Transl. Med. 2014, 12. [Google Scholar] [CrossRef] [PubMed]
- Dufrane, D. Impact of age on human adipose stem cells for bone tissue engineering. Cell Transplant. 2017, 26, 1496–1504. [Google Scholar] [CrossRef] [PubMed]
- Baer, P.C. Adipose-derived mesenchymal stromal/stem cells: An update on their phenotype in vivo and in vitro. World J. Stem Cells 2014, 6, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.; Zevnik, B.; Anastassiadis, K.; Niwa, H.; Klewe-Nebenius, D.; Chambers, I.; Schöler, H.; Smith, A. Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4. Cell 1998, 95, 379–391. [Google Scholar] [CrossRef]
- Tai, M.H.; Chang, C.C.; Kiupel, M.; Webster, J.D.; Olson, L.K.; Trosko, J.E. Oct4 expression in adult human stem cells: Evidence in support of the stem cell theory of carcinogenesis. Carcinogenesis 2005, 26, 495–502. [Google Scholar] [CrossRef]
- Han, S.M.; Han, S.H.; Coh, Y.R.; Jang, G.; Chan Ra, J.; Kang, S.K.; Lee, H.W.; Youn, H.Y. Enhanced proliferation and differentiation of Oct4- and Sox2-overexpressing human adipose tissue mesenchymal stem cells. Exp. Mol. Med. 2014, 46. [Google Scholar] [CrossRef] [Green Version]
- Niwa, H.; Miyazaki, J.-i.; Smith, A.G. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cells. Nat. Genet. 2000, 24, 372–376. [Google Scholar] [CrossRef]
- Zavan, B.; De Francesco, F.; D’Andrea, F.; Ferroni, L.; Gardin, C.; Salzillo, R.; Nicoletti, G.; Ferraro, G.A. Persistence of CD34 stem marker in human lipoma: Searching for cancer stem cells. Int. J. Biol. Sci. 2015, 11, 1127–1139. [Google Scholar] [CrossRef]
- Ranera, B.; Lyahyai, J.; Romero, A.; Vázquez, F.J.; Remacha, A.R.; Bernal, M.L.; Zaragoza, P.; Rodellar, C.; Martín-Burriel, I. Immunophenotype and gene expression profiles of cell surface markers of mesenchymal stem cells derived from equine bone marrow and adipose tissue. Vet. Immunol. Immunopathol. 2011, 144, 147–154. [Google Scholar] [CrossRef]
- Watson, J.E.; Patel, N.A.; Carter, G.; Moor, A.; Patel, R.; Ghansah, T.; Mathur, A.; Murr, M.M.; Bickford, P.; Gould, L.J.; et al. Comparison of markers and functional attributes of human adipose-derived stem cells and dedifferentiated adipocyte cells from subcutaneous fat of an obese diabetic donor. Adv. Wound Care 2014, 3, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Mühlethaler, A.; Bourloud, K.B.; Beck, M.N.; Gross, N. Hypermethylation-mediated regulation of CD44 gene expression in human neuroblastoma. Gene. Chromosom. Cancer 2003, 36, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.S.; Xin, Z.C.; Dai, J.; Lue, T.F. Commonly used mesenchymal stem cell markers and tracking labels: limitations and challenges. Histol. Histopathol. 2013, 28, 1109–1116. [Google Scholar] [CrossRef]
- Chang, H.; Park, S.O.; Jin, U.S.; Hong, K.Y. Characterization of two distinct lipomas: A comparative analysis from surgical perspective. J. Plast. Surg. Hand Surg. 2018, 52, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Varma, M.J.; Breuls, R.G.; Schouten, T.E.; Jurgens, W.J.; Bontkes, H.J.; Schuurhuis, G.J.; van Ham, S.M.; van Milligen, F.J. Phenotypical and functional characterization of freshly isolated adipose tissue-derived stem cells. Stem Cells Dev. 2007, 16, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Taha, M.F.; Javeri, A.; Rohban, S.; Mowla, S.J. Upregulation of pluripotency markers in adipose tissue-derived stem cells by miR-302 and leukemia inhibitory factor. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Franklin, D.M.; Leddy, H.A.; Robey, P.G.; Storms, R.W.; Gimble, J.M. Surface protein characterization of human adipose tissue-derived stromal cells. J. Cell. Physiol. 2001, 189, 54–63. [Google Scholar] [CrossRef]
- Legzdina, D.; Romanauska, A.; Nikulshin, S.; Kozlovska, T.; Berzins, U. Characterization of senescence of culture-expanded human adipose-derived mesenchymal stem cells. Int. J. Stem Cells 2016, 9, 124–136. [Google Scholar] [CrossRef]
- Truong, N.C.; Bui, K.H.T.; Van Pham, P. Characterization of senescence of human adipose-derived stem cells after long-term expansion. In Advances in Experimental Medicine and Biology; Cohen, I.R., Lajtha, A., Lambris, J.D., Paoletti, R., Rezaei, N., Eds.; Springer: New York, NY, USA, 2018; pp. 1–20. ISSN 0065-2598. [Google Scholar]
- Kern, S.; Eichler, H.; Stoeve, J.; Klüter, H.; Bieback, K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef]
- Locke, M.; Windsor, J.; Dunbar, P.R. Human adipose-derived stem cells: Isolation, characterization and applications in surgery. ANZ J. Surg. 2009, 79, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919. [Google Scholar] [CrossRef]
- Sul, H.S. Minireview: Pref-1: Role in adipogenesis and mesenchymal cell fate. Mol. Endocrinol. 2009, 23, 1717–1725. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hudak, C.; Sul, H.S. Role of preadipocyte factor 1 in adipocyte differentiation. Clin. Lipidol. 2010, 5, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Hudak, C.S.; Sul, H.S. Pref-1, a gatekeeper of adipogenesis. Front. Endocrinol. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Mitterberger, M.C.; Lechner, S.; Mattesich, M.; Kaiser, A.; Probst, D.; Wenger, N.; Pierer, G.; Zwerschke, W. DLK1(PREF1) is a negative regulator of adipogenesis in CD105⁺/CD90⁺/CD34⁺/CD31⁻/FABP4⁻ adipose-derived stromal cells from subcutaneous abdominal fat pats of adult women. Stem Cell Res. 2012, 9, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Ciuffi, S.; Zonefrati, R.; Brandi, M.L. Adipose stem cells for bone tissue repair. Clin. Cases Miner. Bone Metab. 2017, 14, 217–226. [Google Scholar] [CrossRef]
- Bhattacharya, I.; Ghayor, C.; Weber, F.E. The use of adipose tissue-derived progenitors in bone tissue engineering—A review. Transfus. Med. Hemother. 2016, 43, 336–343. [Google Scholar] [CrossRef]
- Udagawa, N.; Takahashi, N.; Yasuda, H.; Mizuno, A.; Itoh, K.; Ueno, Y.; Shinki, T.; Gillespie, M.T.; Martin, T.J.; Higashio, K.; et al. Osteoprotegerin produced by osteoblasts is an important regulator in osteoclast development and function. Endocrinology 2000, 141, 3478–3484. [Google Scholar] [CrossRef]
- Yu, H.; de Vos, P.; Ren, Y. Overexpression of osteoprotegerin promotes preosteoblast differentiation to mature osteoblasts. Angle Orthodont. 2011, 81, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Zajdel, A.; Kałucka, M.; Kokoszka-Mikołaj, E.; Wilczok, A. Osteogenic differentiation of human mesenchymal stem cells from adipose tissue and Wharton’s jelly of the umbilical cord. Acta Biochim. Pol. 2017, 64, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Runx2, a multifunctional transcription factor in skeletal development. J. Cell. Biochem. 2002, 87, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of skeletal development by the Runx family of transcription factors. J. Cell. Biochem. 2005, 95, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, T.; Takahashi, N. Regulatory mechanisms of osteoblast and osteoclast differentiation. Oral Dis. 2002, 8, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of bone development and maintenance by Runx2. Front. Biosci. 2008, 13, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Tsao, Y.T.; Huang, Y.J.; Wu, H.H.; Liu, Y.A.; Liu, Y.S.; Lee, O.K. Osteocalcin mediates biomineralization during osteogenic maturation in human mesenchymal stromal cells. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, Z.Y.; Wang, Y.P.; Wu, Z.H.; Yu, B. Dynamic expression profiles of marker genes in osteogenic differentiation of human bone marrow-derived mesenchymal stem cells. Chin. Med. Sci. J. 2015, 30, 108–113. [Google Scholar] [CrossRef]
- Firth, N.A.; Allsobrook, O.; Patel, M. Osteolipoma of the buccal mucosa: A case report. Aust. Dent. J. 2017, 62, 378–381. [Google Scholar] [CrossRef]
- Demiralp, B.; Alderete, J.F.; Kose, O.; Ozcan, A.; Cicek, I.; Basbozkurt, M. Osteolipoma independent of bone tissue: A case report. Cases J. 2009, 2. [Google Scholar] [CrossRef]
- Raghunath, V.; Manjunatha, B.S. Osteolipoma of floor of the mouth. BMJ Case Rep. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Kwan Ip, N.S.; Lau, H.W.; Wong, W.Y.; Yuen, M.K. Osteolipoma in the Forearm. J. Clin. Imaging Sci. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, H.; Furuichi, T.; Zanma, A.; Yamana, K.; Yoshida, C.; Sumitani, S.; Yamamoto, H.; Enomoto-Iwamoto, M.; Iwamoto, M.; Komori, T. Runx2 deficiency in chondrocytes causes adipogenic changes in vitro. J. Cell Sci. 2004, 117, 417–425. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
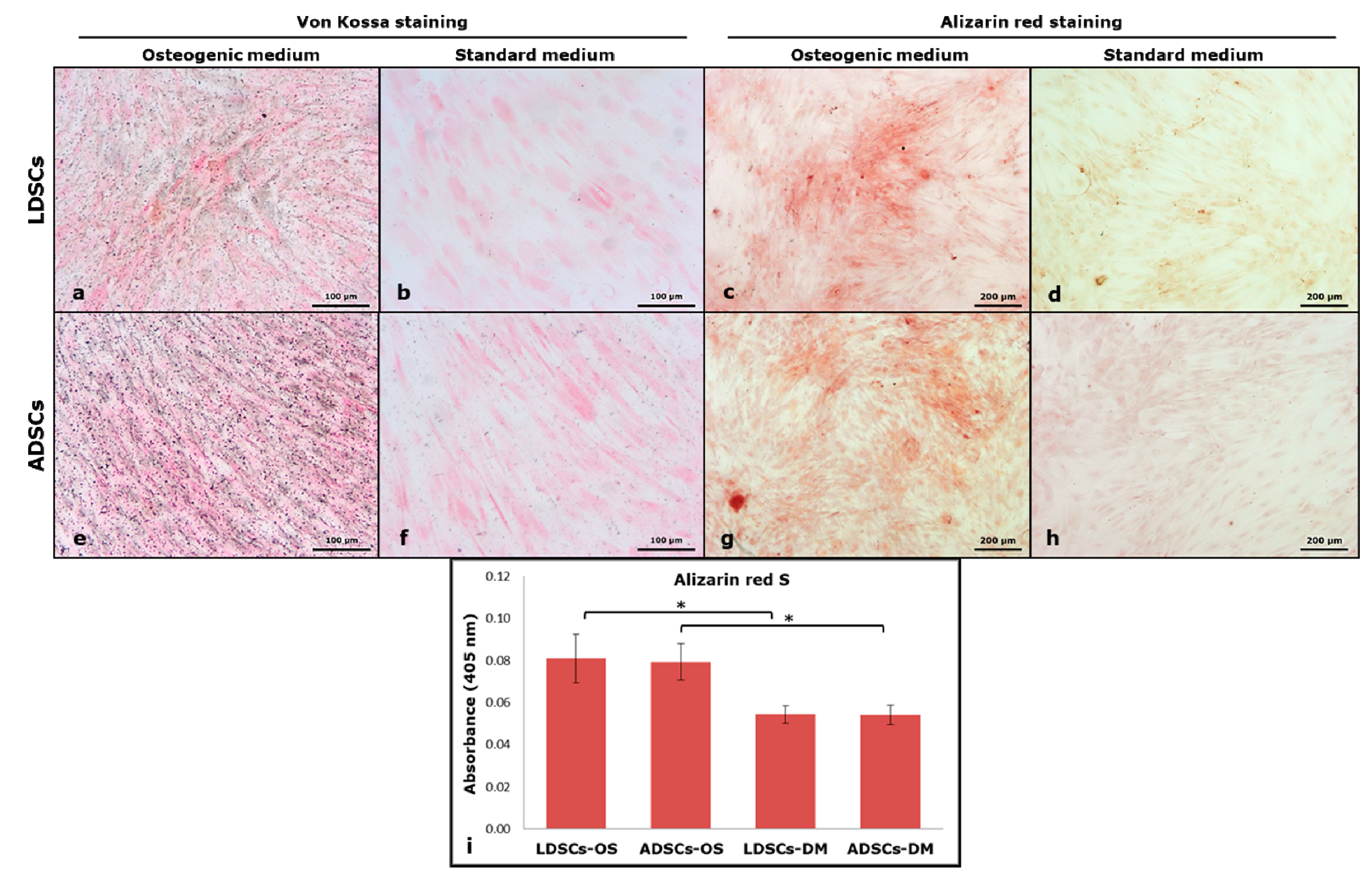{kind=link}
{kind=link}
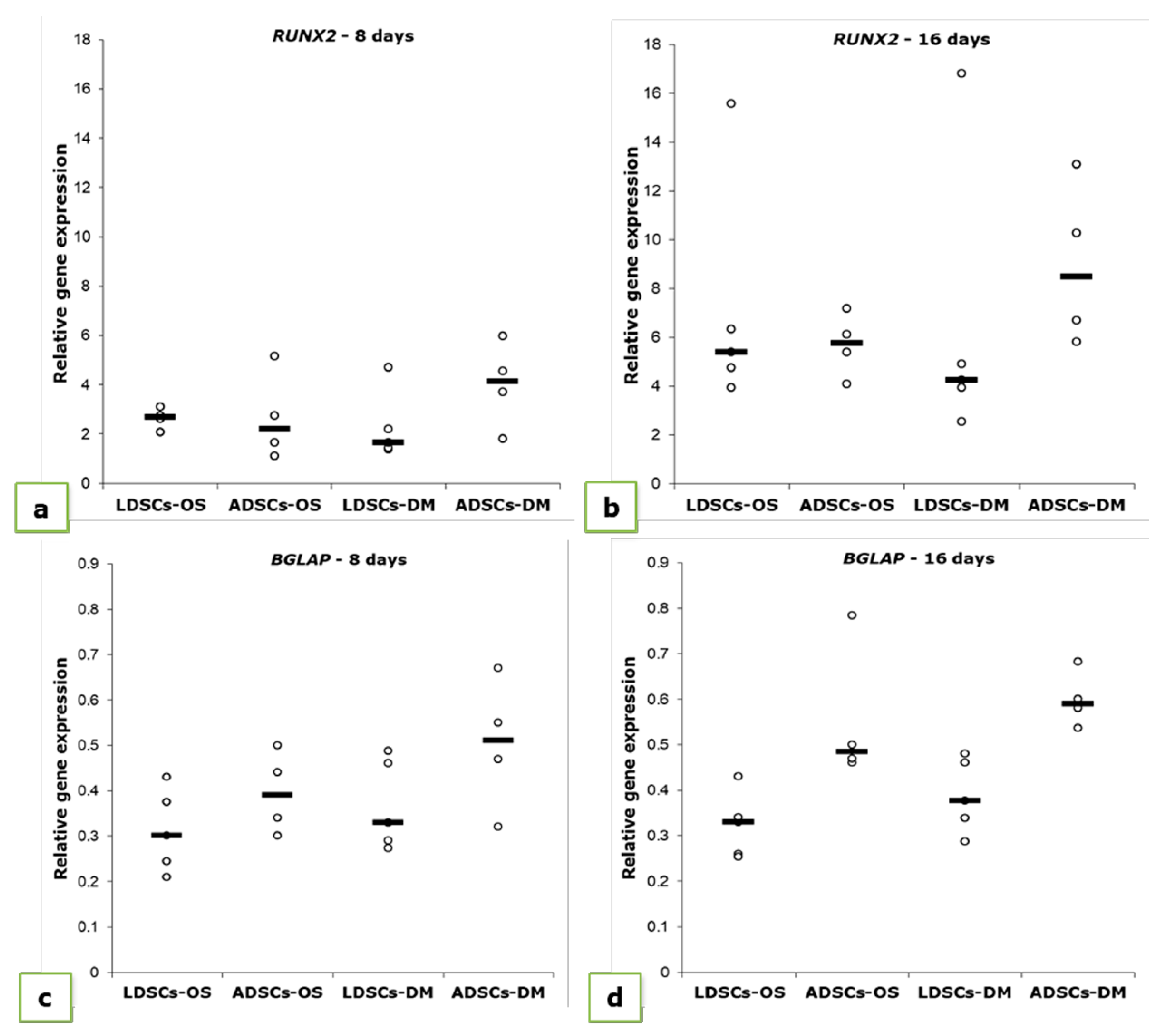{kind=link}
| Cell Type | ADIPOQ | LEP | ||||
|---|---|---|---|---|---|---|
| P2 (Day 0) | 21 Day | P2 (Day 0) | 21 Day | |||
| AM | DM | AM | DM | |||
| LDSCs | 0.033 ± 0.033 | 12.530 ± 7.31 * | 0.400 ± 0.35 | 0.0130 ± 0.011 | 0.03 ± 0.012 * | 0.030 ± 0.025 |
| ADSCs | 0.061 ± 0.019 | 993.84 ± 174.09 *** | 0.515 ± 0.45 | 0.0041 ± 0.003 | 0.46 ± 0.41 *** | 0.025 ± 0.006 ** |
| PPARG | DLK1 | |||||
| P2 (Day 0) | 21 Day | P2 (Day 0) | 21 Day | |||
| AM | DM | AM | DM | |||
| LDSCs | 0.159 ± 0.151 | 0.30 ± 0.187 | 0.32 ± 0.247 | 0.32 ± 0.725 | 6.86 ± 4.38 * | 1.50 ± 1.73 |
| ADSCs | 0.250 ± 0.188 | 4.92 ± 3.50 * | 0.42 ± 0.315 | 0.10 ± 0.105 | 0.57 ± 1.13 | 2.61 ± 4.92 |
| Cell Type | RUNX2 | ||||
|---|---|---|---|---|---|
| P2 (Day 0) | 8 Days | 16 Days | |||
| OS | DM | OS | DM | ||
| LDSCs | 1.620 ± 0.622 | 2.156 ± 1.133 | 2.27 ± 1.390 | 7.194 ± 4.76 * | 6.478 ± 5.83 |
| ADSCs | 1.013 ± 0.214 | 2.670 ± 1.799 | 4.01 ± 1.747 * | 5.685 ± 1.29 *** | 8.960 ± 3.35 ** |
| BGLAP | |||||
| P2 (Day 0) | 8 Days | 16 Days | |||
| OS | DM | OS | DM | ||
| LDSCs | 0.23 ± 0.039 | 0.312 ± 0.091 * | 0.368 ± 0.099 * | 0.323 ± 0.072 * | 0.388 ± 0.081 ** |
| ADSCs | 0.33 ± 0.085 | 0.395 ± 0.091 | 0.503 ± 0.147 | 0.554 ± 0.155 * | 0.600 ± 0.062 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stojanović, S.; Najman, S.; Korać, A. Stem Cells Derived from Lipoma and Adipose Tissue—Similar Mesenchymal Phenotype but Different Differentiation Capacity Governed by Distinct Molecular Signature. Cells 2018, 7, 260. https://doi.org/10.3390/cells7120260
Stojanović S, Najman S, Korać A. Stem Cells Derived from Lipoma and Adipose Tissue—Similar Mesenchymal Phenotype but Different Differentiation Capacity Governed by Distinct Molecular Signature. Cells. 2018; 7(12):260. https://doi.org/10.3390/cells7120260
Chicago/Turabian StyleStojanović, Sanja, Stevo Najman, and Aleksandra Korać. 2018. "Stem Cells Derived from Lipoma and Adipose Tissue—Similar Mesenchymal Phenotype but Different Differentiation Capacity Governed by Distinct Molecular Signature" Cells 7, no. 12: 260. https://doi.org/10.3390/cells7120260