Transected Tendon Treated with a New Fibrin Sealant Alone or Associated with Adipose-Derived Stem Cells

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of ASC and Ccell Culture
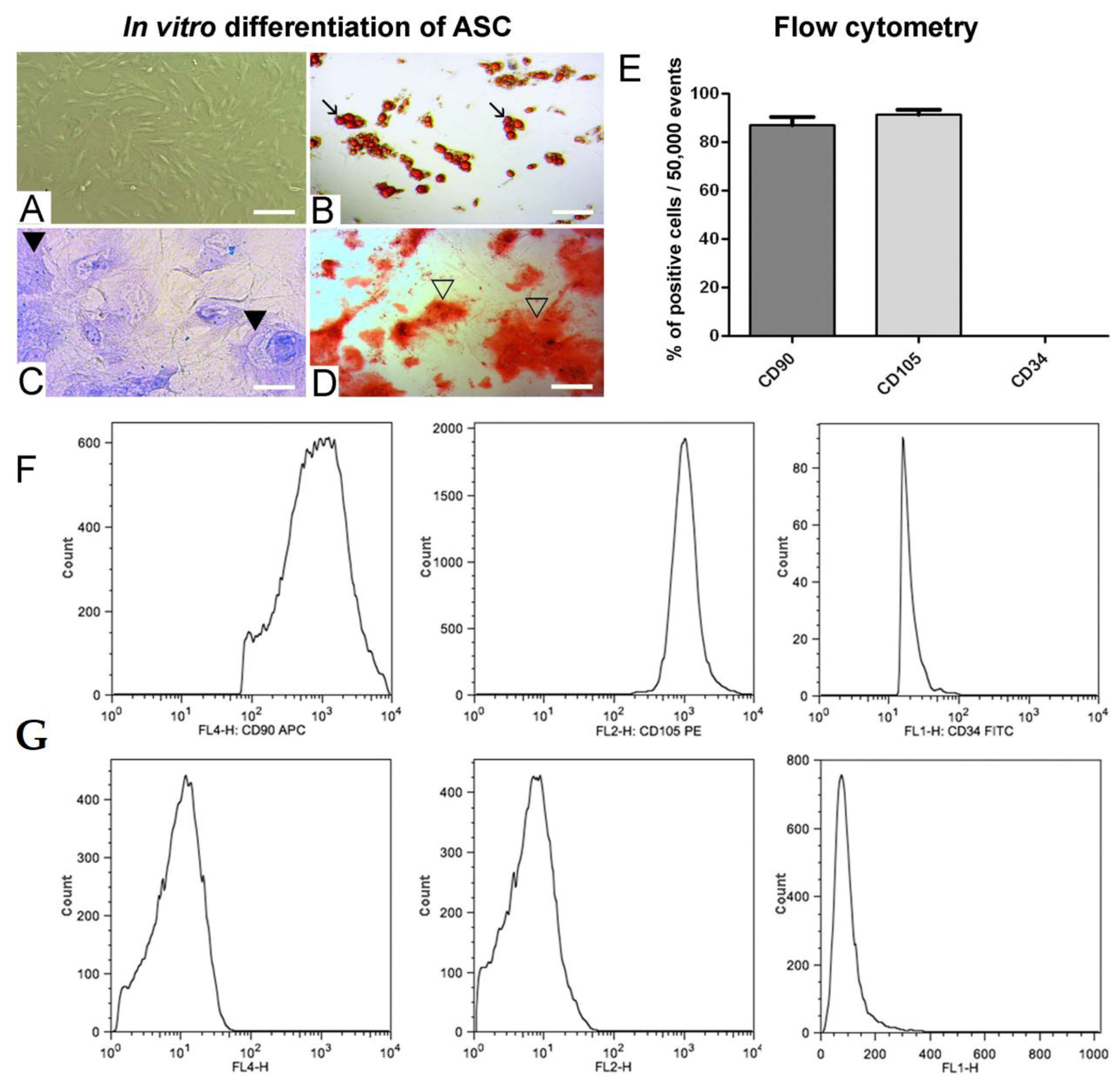
2.2. Flow Cytometry
2.3. In Vitro Differentiation Potential of ASC
2.4. Fibrin Sealant (FS) Scaffold
2.5. Confocal Microscope Analysis
2.6. Experimental Groups
2.7. Protocol for Partial Transection of the Achilles Tendon and Application of FS and ASC
2.8. In Vivo Imaging
2.9. Real-Time PCR Array
2.10. Dosage of Hydroxyproline
2.11. Western Blotting
2.12. Preparation of Sections in Freezing
2.13. Birefringence and Contrast by Differential Interference (DIC)
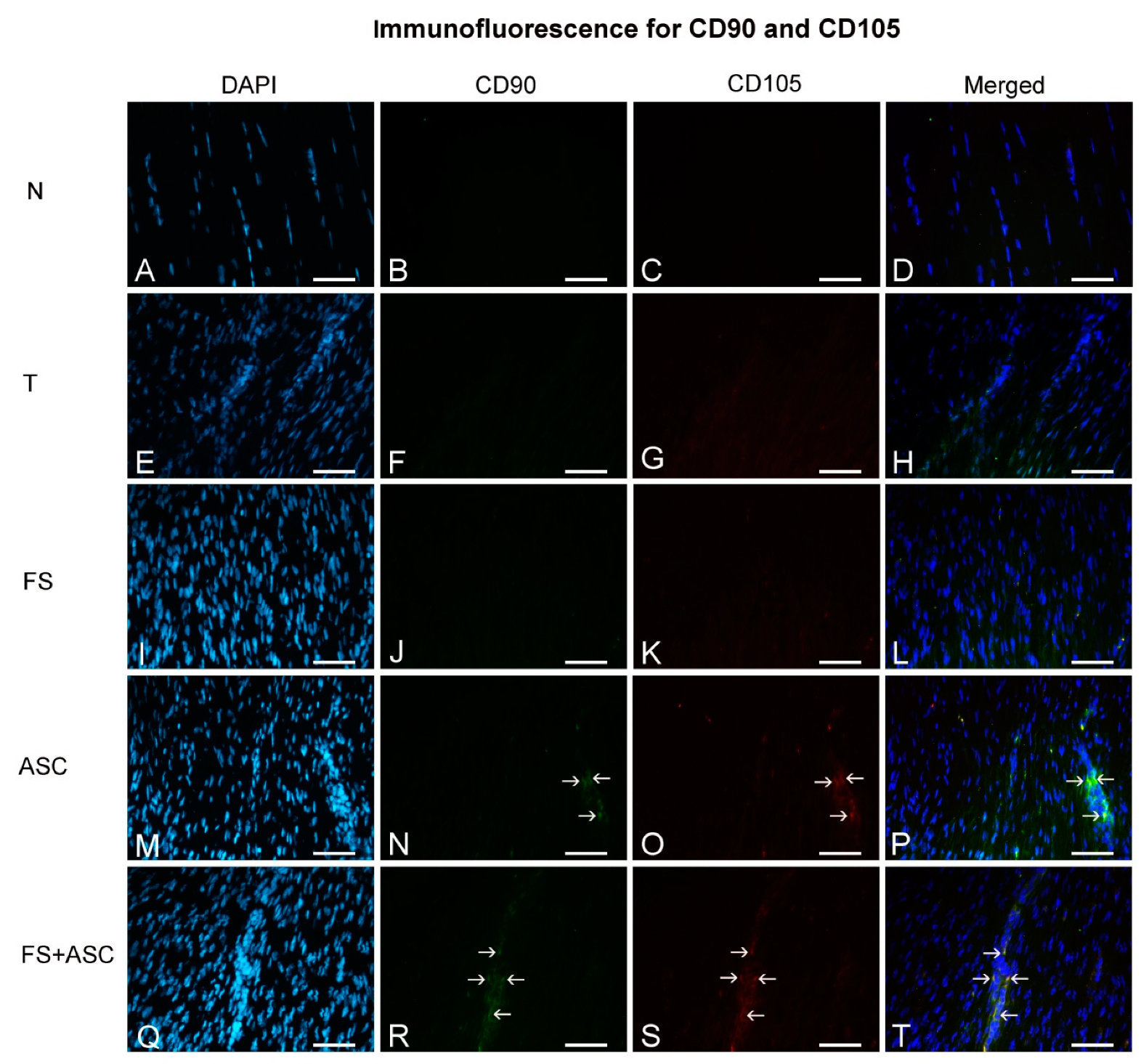
2.14. Immunofluorescence
2.15. Histology and Histomorphometry
2.16. Biomechanical Parameters
2.17. Statistical Analysis
3. Results
3.1. In Vitro Adipogenic, Chondrogenic and Osteogenic Differentiation of ASC and Positive ASC Markers
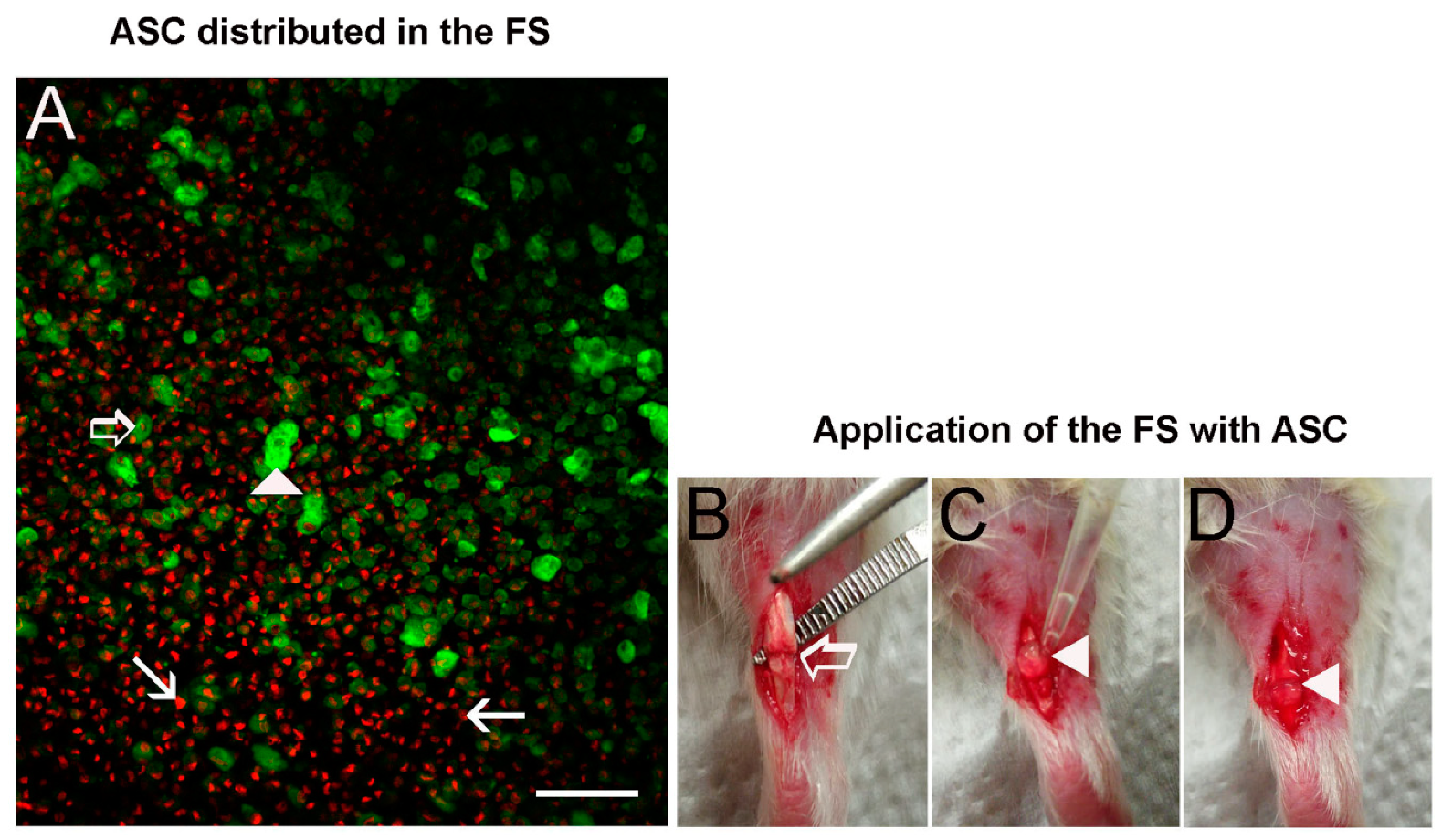
3.2. ASC Disposition in the FS and Application on the Transected Tendon
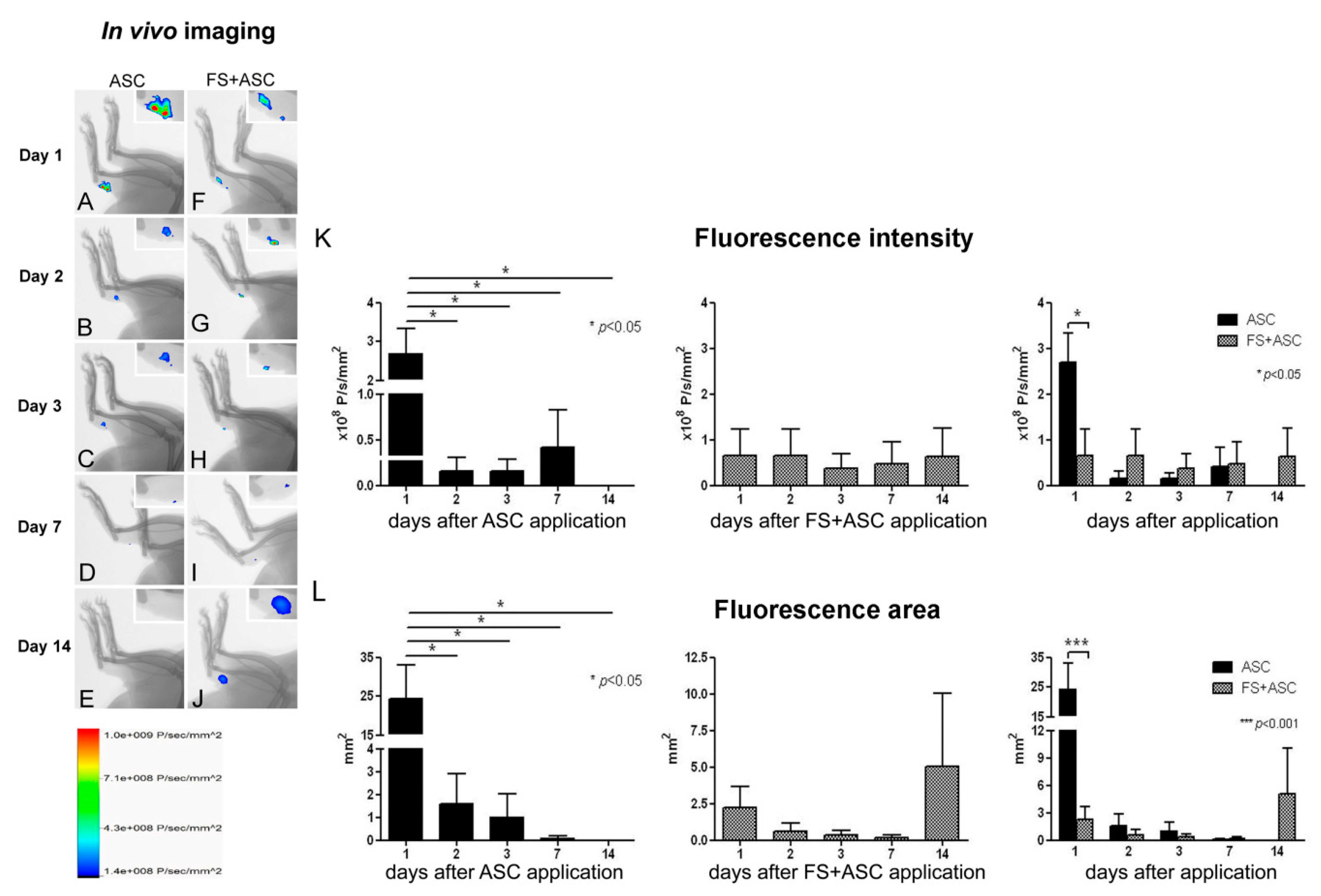
3.3. In Vivo Imaging for ASC Detection on Tendon
3.4. Cell Migration Assay and Macrophages Identification
3.5. Genes Expression Analysis
3.6. Total Collagen Quantification
3.7. Collagen I and Tnmd quantification
3.8. Panoramic View of Tendons Organization and Histomorphometry
3.9. Collagen Fibers Organization Measurements
3.10. Biomechanical Properties of Tendons
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nourissat, G.; Berenbaum, F.; Duprez, D. Tendon injury: From biology to tendon repair. Nat. Rev. Rheumatol. 2015, 11, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Andarawis-Puri, N.; Flatow, E.L.; Soslowsky, L.J. Tendon Basic Science: Development, Repair, Regeneration, and Healing. J. Orthop. Res. 2015, 33, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, D.L.; Huang, E.K.; Silver, F.H. Assembly of type I collagen: Fusion of fibril subunits and the influence of fibril diameter on mechanical properties. Matrix Biol. 2000, 19, 409–420. [Google Scholar] [CrossRef]
- Vidal, B.C. Cell and extracellular matrix interaction: A feedback theory based on molecular order recognition-adhesion events. Rev. Fac. Ciên. Med. Unicamp. 1994, 4, 11–14. [Google Scholar]
- Lavagnino, M.; Wall, M.E.; Little, D.; Banes, A.J.; Guilak, F.; Arnoczky, S.P. Tendon Mechanobiology: Current Knowledge and Future Research Opportunities. J. Orthop. Res. 2015, 33, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Arnoczky, S.P.; Lavagnino, M.; Whallon, J.H.; Hoonjan, A. In situ cell nucleus deformation in tendons under tensile load; a morphological analysis using confocal laser microscopy. J. Orthop. Res. 2002, 20, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Arnoczky, S.P.; Lavagnino, M.; Egerbacher, M. The mechanobiological aetiopathogenesis of tendinopathy: Is it the over-stimulation or the under-stimulation of tendon cells? Int. J. Exp. Pathol. 2007, 88, 217–226. [Google Scholar] [CrossRef]
- Archambault, J.M.; Wiley, J.P.; Bray, R.C. Exercise loading of tendons and the development of overuse injuries. A review of current literature. Sports Med. 1995, 20, 77–89. [Google Scholar] [CrossRef]
- Durgam, S.; Stewart, M. Cellular and Molecular Factors Influencing Tendon Repair. Tissue Eng. Part B Rev. 2017, 23, 307–317. [Google Scholar] [CrossRef]
- Docheva, D.; Müller, S.A.; Majewski, M.; Evans, C.H. Biologics for tendon repair. Adv. Drug Deliv. Rev. 2015, 84, 222–239. [Google Scholar] [CrossRef] [Green Version]
- Aro, A.A.; Nishan, U.; Perez, M.O.; Rodrigues, R.A.; Foglio, M.A.; Carvalho, J.E.; Gomes, L.; Vidal, B.C.; Pimentel, E.R. Structural and biochemical alterations during the healing process of tendons treated with Aloe vera. Life Sci. 2012, 91, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Leadbetter, W.B. Cell-matrix response in tendon injury. Clin. Sports Med. 1992, 11, 533–578. [Google Scholar] [PubMed]
- Merceron, C.; Vinatier, C.; Clouet, J.; Colliec-Jouault, S.; Weiss, P.; Guicheux, J. Adipose-derived mesenchymal stem cells and biomaterials for cartilage tissue engineering. Jt. Bone Spine 2008, 75, 672–674. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.H.; Rousche, K.T.; Tuan, R.S. Technology Insight: Adult stem cells in cartilage regeneration and tissue engineering. Nat. Clin. Pract. Rheumatol. 2006, 2, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.I.; Rodrigues, M.T.; Lee, S.J.; Atala, A.; Yoo, J.J.; Reis, R.L.; Gomes, M.E. Understanding the Role of Growth Factors in Modulating Stem Cell Tenogenesis. PLoS ONE 2013, 8, e83734. [Google Scholar] [CrossRef] [PubMed]
- Oedayrajsingh-Varma, M.J.; Van Ham, S.M.; Knippenberg, M.; Helder, M.N.; Klein-Nulend, J.; Schouten, T.E.; Ritt, M.J.; van Milligen, F.J. Adipose tissue-derived mesenchymal stem cell yield and growth characteristics are affected by the tissue-harvesting procedure. Cytotherapy 2006, 8, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Katz, A.J.; Tholpady, A.; Tholpady, S.S.; Shang, H.; Ogle, R.C. Cell surface and transcriptional characterization of human adipose-derived adherent stromal (hADAS) cells. Stem Cells 2005, 23, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Ling, W.; Xu, W.; Han, D. Fibroblast-Like Cells Differentiated from Adipose-Derived Mesenchymal Stem Cells for Vocal Fold Wound Healing. PLoS ONE 2014, 9, e92676. [Google Scholar] [CrossRef]
- Altman, A.M.; Matthias, N.; Yan, Y.; Song, Y.-H.; Bai, X.; Chiu, E.S.; Slakey, D.P.; Alt, E.U. Dermal matrix as a carrier for in vivo delivery of human adipose-derived stem cells. Biomaterials 2008, 29, 1431–1442. [Google Scholar] [CrossRef]
- Kern, S.; Eichler, H.; Stoeve, J.; Klüter, H.; Bieback, K. Comparative Analysis of Mesenchymal Stem Cells from Bone Marrow, Umbilical Cord Blood, or Adipose Tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.-M.; Elabd, C.; Amri, E.-Z.; Ailhaud, G.; Dani, C. The human adipose tissue is a source of multipotent stem cells. Biochimie 2005, 87, 125–128. [Google Scholar] [CrossRef] [PubMed]
- De Ugarte, D.A.; Morizono, K.; Elbarbary, A.; Alfonso, Z.; Zuk, P.A.; Zhu, M.; Dragoo, J.L.; Ashjian, P.; Thomas, B.; Benhaim, P.; et al. Comparison of Multi-Lineage Cells from Human Adipose Tissue and Bone Marrow. Cells Tissues Organs 2003, 174, 101–109. [Google Scholar] [CrossRef]
- Hassan, W.U.; Greiser, U.; Wang, W. Role of adipose-derived stem cells in wound healing. Wound Repair Regen. 2014, 22, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Johnstone, B.H.; Cook, T.G.; Liang, Z.; Traktuev, D.; Cornetta, K.; Ingram, D.A.; Rosen, E.D.; March, K.L. Suppression of hepatocyte growth factor production impairs the ability of adipose derived stem cells to promote ischemic tissue revascularization. Stem Cells 2007, 25, 3234–3243. [Google Scholar] [CrossRef] [PubMed]
- Hattori, H.; Ishihara, M. Altered protein secretions during interactions between adipose tissue- or bone marrow-derived stromal cells and inflammatory cells. Stem Cell Res. Ther. 2015, 6, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uysal, C.A.; Tobita, M.; Hyakusoku, H.; Mizuno, H. Adipose-derived stem cells enhance primary tendon repair: Biomechanical and immunohistochemical evaluation. J. Plast. Reconstr. Aesthet. Surg. 2012, 65, 1712–1719. [Google Scholar] [CrossRef]
- Kim, J.H.; Jung, M.; Kim, H.S.; Kim, Y.M.; Choi, E.H. Adipose-derived stem cells as a new therapeutic modality for ageing skin. Exp. Dermatol. 2011, 20, 383–387. [Google Scholar] [CrossRef]
- Shingyochi, Y.; Orbay, H.; Mizuno, H. Adipose-derived stem cells for wound repair and regeneration. Expert Opin. Biol. Ther. 2015, 15, 1285–1292. [Google Scholar] [CrossRef]
- Gasparotto, V.P.O.; Landim-Alvarenga, F.C.; Oliveira, A.L.R.; Simões, G.; Lima-Neto, J.F.; Barraviera, B.; Ferreira, R.S. A new fibrin sealant as a three-dimensional scaffold candidate for mesenchymal stem cells. Stem Cell Res. Ther. 2014, 5, 78. [Google Scholar] [CrossRef] [Green Version]
- James, R.; Kumbar, S.G.; Laurencin, C.T.; Balian, G.; Chhabra, A.B. Tendon tissue engineering: Adipose-derived stem cell and GDF-5 mediated regeneration using electrospun matrix systems. Biomed. Mater. 2011, 6, 025011. [Google Scholar] [CrossRef]
- Biscola, N.P.; Cartarozzi, L.P.; Ulian-Benitez, S.; Barbizan, R.; Castro, M.V.; Spejo, A.B.; Ferreira, R.S.; Barraviera, B.; Oliveira, A.L.R. Multiple uses of fibrin sealant for nervous system treatment following injury and disease. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 13. [Google Scholar] [CrossRef] [PubMed]
- Thomazini-Santos, I.A. Fibrin adhesive from snake venom: The effect of adding epsilon-aminocaproic acid, tranexamic acid, and aprotinin for coaptation of wound edges in rat skin incisions. J. Venom. Anim. Toxins 2001, 7, 148–149. [Google Scholar] [CrossRef]
- Thomazini-Santos, I.A.; Barraviera, S.R.C.S.; Mendes-Giannini, M.J.S.; Barraviera, B. Surgical adhesives. J. Venom. Anim. Toxins 2001, 7, 159–171. [Google Scholar] [CrossRef]
- Ferreira, R.S.; de Barros, L.C.; Abbade, L.P.F.; Barraviera, S.R.C.S.; Silvares, M.R.C.; de Pontes, L.G.; dos Santos, L.D.; Barraviera, B. Heterologous fibrin sealant derived from snake venom: From bench to bedside—An overview. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 21. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.C.; Ferreira, R.S.; Barraviera, S.R.C.S.; Stolf, H.O.; Thomazini-Santos, I.A.; Mendes-Giannini, M.J.S.; Toscano, E.; Barraviera, B. A new fibrin sealant from Crotalus durissus terrificus venom: Applications in medicine. J. Toxicol. Environ. Health Part B Crit. Rev. 2009, 12, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Gazzeri, R.; Fiore, C.; Galarza, M. Role of EVICEL Fibrin Sealant to Assist Hemostasis in Cranial and Spinal Epidural Space: A Neurosurgical Clinical Study. Surg. Technol. Int. 2015, 26, 364–369. [Google Scholar] [PubMed]
- Scognamiglio, F.; Travan, A.; Rustighi, I.; Tarchi, P.; Palmisano, S.; Marsich, E.; Borgogna, M.; Donati, I.; de Manzini, N.; Paoletti, S. Adhesive and sealant interfaces for general surgery applications. J. Biomed. Mater. Res. Part B Appl. Biomater. 2016, 104, 626–639. [Google Scholar] [CrossRef]
- Noori, A.; Ashrafi, S.J.; Vaez-Ghaemi, R.; Hatamian-Zaremi, A.; Webster, T.J. A review of fibrin and fibrin composites for bone tissue engineering. Int. J. Nanomed. 2017, 12, 4937–4961. [Google Scholar] [CrossRef] [Green Version]
- Barsotti, M.C.; Felice, F.; Balbarini, A.; Di Stefano, R. Fibrin as a scaffold for cardiac tissue engineering. Biotechnol. Appl. Biochem. 2011, 58, 301–310. [Google Scholar] [CrossRef]
- Sameem, M.; Wood, T.J.; Bain, J.R. A systematic review on the use of fibrin glue for peripheral nerve repair. Plast Reconstr. Surg. 2011, 127, 2381–2390. [Google Scholar] [CrossRef]
- Kayaalp, C.; Ertugrul, I.; Tolan, K.; Sumer, F. Fibrin sealant use in pilonidal sinus: Systematic review. World J. Gastrointest. Surg. 2016, 8, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-F.; He, X.; He, J.; Zhang, L.-H.; Su, X.-J.; Dong, Z.-Y.; Xu, Y.-J.; Li, Y.; Li, Y.-L. High efficient isolation and systematic identification of human adipose-derived mesenchymal stem cells. J. Biomed. Sci. 2011, 18, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.S. Autologous or heterologous fibrin sealant scaffold: Which is the better choice? J. Venom. Anim. Toxins Incl. Trop. Dis. 2014, 20, 31. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Soares, A.; Costa, F.; Rodrigues, V.; Fuly, A.; Giglio, J.; Gallacci, M.; Thomazini-Santos, I.; Barraviera, S.; Barraviera, B.; et al. Biochemical and biological evaluation of gyroxin isolated from Crotalus durissus terrificus venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2011, 17, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Aro, A.A.; Perez, M.O.; Vieira, C.P.; Esquisatto, M.A.M.; Rodrigues, R.A.F.; Gomes, L.; Pimentel, E.R. Effect of calendula officinalis cream on achilles tendon healing. Anat. Rec. 2015, 298, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Aro, A.A.; Esquisatto, M.A.M.; Nishan, U.; Perez, M.O.; Rodrigues, R.A.F.; Foglio, M.A.; De Carvalho, J.E.; Gomes, L.; Vidal, B.D.C.; Pimentel, E.R. Effect of A loe vera application on the content and molecular arrangement of glycosaminoglycans during calcaneal tendon healing. Microsc. Res. Tech. 2014, 77, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Aro, A.A.; Freitas, K.M.; Foglio, M.A.; Carvalho, J.E.; Dolder, H.; Gomes, L.; Vidal, B.C.; Pimentel, E.R. Effect of the Arrabidaea chica extract on collagen fiber organization during healing of partially transected tendon. Life Sci. 2013, 92, 799–807. [Google Scholar] [CrossRef] [Green Version]
- Aro, A.A.; Simões, G.F.; Esquisatto, M.A.M.; Foglio, M.A.; Carvalho, J.E.; Oliveira, A.L.R.; Gomes, L.; Pimentel, E.R. Arrabidaea chica extract improves gait recovery and changes collagen content during healing of the Achilles tendon. Injury 2013, 44, 884–892. [Google Scholar] [CrossRef]
- Guerra, F.D.R.; Vieira, C.P.; Almeida, M.S.; Oliveira, L.P.; De Aro, A.A.; Pimentel, E.R. LLLT improves tendon healing through increase of MMP activity and collagen synthesis. Lasers Med. Sci. 2013, 28, 1281–1288. [Google Scholar] [CrossRef]
- Tomiosso, T.C.; Nakagaki, W.R.; Gomes, L.; Hyslop, S.; Pimentel, E.R. Organization of collagen bundles during tendon healing in rats treated with L-NAME. Cell Tissue Res. 2009, 337, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Godoy, J.A.P.; Block, D.B.; Tollefsen, D.M.; Werneck, C.C.; Vicente, C.P. Dermatan sulfate and bone marrow mononuclear cells used as a new therapeutic strategy after arterial injury in mice. Cytotherapy 2011, 13, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Marqueti, R.C.; Durigan, J.L.Q.; Oliveira, A.J.S.; Mekaro, M.S.; Guzzoni, V.; Aro, A.A.; Pimentel, E.R.; Selistre-De-Araujo, H.S. Effects of aging and resistance training in rat tendon remodeling. FASEB J. 2018, 32, 353–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stegemann, H.; Stalder, K. Determination of hydroxyproline. Clin. Chim. Acta 1967, 18, 267–273. [Google Scholar] [CrossRef]
- Jorge, M.P.; Madjarof, C.; Ruiz, A.L.T.G.; Fernandes, A.T.; Rodrigues, R.A.F.; de Oliveira Sousa, I.M.; Foglio, M.A.; de Carvalho, J.E. Evaluation of wound healing properties of Arrabidaea chica Verlot extract. J. Ethnopharmacol. 2008, 118, 361–366. [Google Scholar] [CrossRef] [PubMed]
- de Campos Vidal, B. Fluorescence, aggregation properties and FT-IR microspectroscopy of elastin and collagen fibers. Acta Histochem. 2014, 116, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- de Campos Vidal, B.; dos Anjos, E.H.M.; Mello, M.L.S. Optical anisotropy reveals molecular order in a mouse enthesis. Cell Tissue Res. 2015, 362, 177–185. [Google Scholar] [CrossRef]
- de Campos Vidal, B. Form birefringence as applied to biopolymer and inorganic material supraorganization. Biotech. Histochem. 2010, 85, 365–378. [Google Scholar] [CrossRef]
- de Campos Vidal, B.; Mello, M.L.S. Optical anisotropy of collagen fibers of rat calcaneal tendons: An approach to spatially resolved supramolecular organization. Acta Histochem. 2010, 112, 53–61. [Google Scholar] [CrossRef]
- Biancalana, A.; Veloso, L.; Gomes, L. Obesity affects collagen fibril diameter and mechanical properties of tendons in Zucker rats. Connect. Tissue Res. 2010, 51, 171–178. [Google Scholar] [CrossRef]
- Sparrow, K.J.; Finucane, S.D.; Owen, J.R.; Wayne, J.S. The Effects of Low-Intensity Ultrasound on Medial Collateral Ligament Healing in the Rabbit Model. Am. J. Sports Med. 2005, 33, 1048–1056. [Google Scholar] [CrossRef]
- Qiao, H.; Surti, S.; Choi, S.R.; Raju, K.; Zhang, H.; Ponde, D.E.; Kung, H.F.; Karp, J.; Zhou, R. Death and proliferation time course of stem cells transplanted in the myocardium. Mol. Imaging Biol. 2009, 11, 408–414. [Google Scholar] [CrossRef]
- Wolbank, S.; Pichler, V.; Ferguson, J.C.; Meinl, A.; Van Griensven, M.; Goppelt, A.; Redl, H. Non-invasive in vivo tracking of fibrin degradation by fluorescence imaging. J. Tissue Eng. Regen. Med. 2015, 9, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Bensaïd, W.; Triffitt, J.T.; Blanchat, C.; Oudina, K.; Sedel, L.; Petite, H. A biodegradable fibrin scaffold for mesenchymal stem cell transplantation. Biomaterials 2003, 24, 2497–2502. [Google Scholar] [CrossRef]
- Spejo, A.B.; Chiarotto, G.B.; Ferreira, A.D.F.; Gomes, D.A.; Ferreira, R.S., Jr.; Barraviera, B.; Oliveira, A.L.R. Neuroprotection and immunomodulation following intraspinal axotomy of motoneurons by treatment with adult mesenchymal stem cells. J. Neuroinflamm. 2018, 15, 230. [Google Scholar] [CrossRef] [PubMed]
- Orsi, P.R.; Landim-Alvarenga, F.C.; Justulin, L.A., Jr.; Kaneno, R.; Golim, M.A.; Santos, D.C.; Creste, C.F.Z.; Oba, E.; Maia, L.; Barraviera, B.; et al. A unique heterologous fibrin sealant (HFS) as a candidate biological scaffold for mesenchymal stem cells in osteoporotic rats. Stem Cell Res. Ther. 2017, 8, 205. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Cao, W.; Shi, Y. Plasticity of mesenchymal stem cells in immunomodulation: pathological and therapeutic implications. Nat. Immunol. 2014, 15, 1009–1016. [Google Scholar] [CrossRef]
- Peroni, J.F.; Borjesson, D.L. Anti-Inflammatory and Immunomodulatory Activities of Stem Cells. Vet. Clin. N. Am. Equine Pract. 2011, 27, 351–362. [Google Scholar] [CrossRef]
- Lee, R.H.; Oh, J.Y.; Choi, H.; Bazhanov, N. Therapeutic factors secreted by mesenchymal stromal cells and tissue repair. J. Cell. Biochem. 2011, 112, 3073–3078. [Google Scholar] [CrossRef]
- Veronesi, F.; Salamanna, F.; Tschon, M.; Maglio, M.; Nicoli Aldini, N.; Fini, M. Mesenchymal stem cells for tendon healing: What is on the horizon? J. Tissue Eng. Regen. Med. 2017, 11, 3202–3219. [Google Scholar] [CrossRef]
- Raabe, O.; Shell, K.; Fietz, D.; Freitag, C.; Ohrndorf, A.; Christ, H.J.; Wenisch, S.; Arnhold, S. Tenogenic differentiation of equine adipose-tissue-derived stem cells under the influence of tensile strain, growth differentiation factors and various oxygen tensions. Cell Tissue Res. 2013, 352, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Shukunami, C.; Takimoto, A.; Oro, M.; Hiraki, Y. Scleraxis positively regulates the expression of tenomodulin, a differentiation marker of tenocytes. Dev. Biol. 2006, 298, 234–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lui, P.P.Y. Identity of tendon stem cells—How much do we know? J. Cell. Mol. Med. 2013, 17, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Uysal, A.C.; Mizuno, H. Differentiation of Adipose-Derived Stem Cells for Tendon Repair. In Methods in Molecular Biology (Clifton, N.J.); Humana Press: Totowa, NJ, USA, 2011; Volume 702, pp. 443–451. [Google Scholar]
- James, R.; Kesturu, G.; Balian, G.; Chhabra, A.B. Tendon: Biology, Biomechanics, Repair, Growth Factors, and Evolving Treatment Options. J. Hand Surg. Am. 2008, 33, 102–112. [Google Scholar] [CrossRef]
- Kyurkchiev, D.; Bochev, I.; Ivanova-Todorova, E.; Mourdjeva, M.; Oreshkova, T.; Belemezova, K.; Kyurkchiev, S. Secretion of immunoregulatory cytokines by mesenchymal stem cells. World J. Stem Cells 2014, 6, 552. [Google Scholar] [CrossRef]
- Klein, M.B.; Yalamanchi, N.; Pham, H.; Longaker, M.T.; Chang, J. Flexor tendon healing in vitro: Effects of TGF-beta on tendon cell collagen production. J. Hand Surg. Am. 2002, 27, 615–620. [Google Scholar] [CrossRef]
- Rickert, M.; Jung, M.; Adiyaman, M.; Richter, W.; Simank, H.G. A growth and differentiation factor-5 (GDF-5)-coated suture stimulates tendon healing in an Achilles tendon model in rats. Growth Factors 2001, 19, 115–126. [Google Scholar] [CrossRef]
- Peres, V.S. [Unesp] Efeito Do Selante De Fibrina Derivado De Peçonha De Serpente Associado A Células—Tronco Mesenquimais Na Cicatrização De Ferida Cirúrgica em Ratos. Dissertação de Mestrado UNESP, Universidade Estadual Paulista, Sao Paulo, Brazil, 2014. [Google Scholar]
- Yücel, E.A.; Oral, O.; Olgaç, V.; Oral, C.K. Effects of fibrin glue on wound healing in oral cavity. J. Dent. 2003, 31, 569–575. [Google Scholar] [CrossRef]
- Chailakhyan, R.K.; Shekhter, A.B.; Ivannikov, S.V.; Tel’pukhov, V.I.; Suslin, D.S.; Gerasimov, Y.V.; Tonenkov, A.M.; Grosheva, A.G.; Panyushkin, P.V.; Moskvina, I.L.; et al. Reconstruction of Ligament and Tendon Defects Using Cell Technologies. Bull. Exp. Biol. Med. 2017, 162, 563–568. [Google Scholar] [CrossRef]
- Tuan, T.-L.; Song, A.; Chang, S.; Younai, S.; Nimni, M.E. In VitroFibroplasia: Matrix Contraction, Cell Growth, and Collagen Production of Fibroblasts Cultured in Fibrin Gels. Exp. Cell Res. 1996, 223, 127–134. [Google Scholar] [CrossRef]
- Ferraro, G.C.; Moraes, J.R.; Shimano, A.C.; Pereira, G.T.; Moraes, F.R.; Bueno de Camargo, M.H. Effect of snake venom derived fibrin glue on the tendon healing in dogs: Clinical and biomechanical study. J. Venom. Anim. Toxins Incl. Trop. Dis. 2005, 11, 261–274. [Google Scholar] [CrossRef]




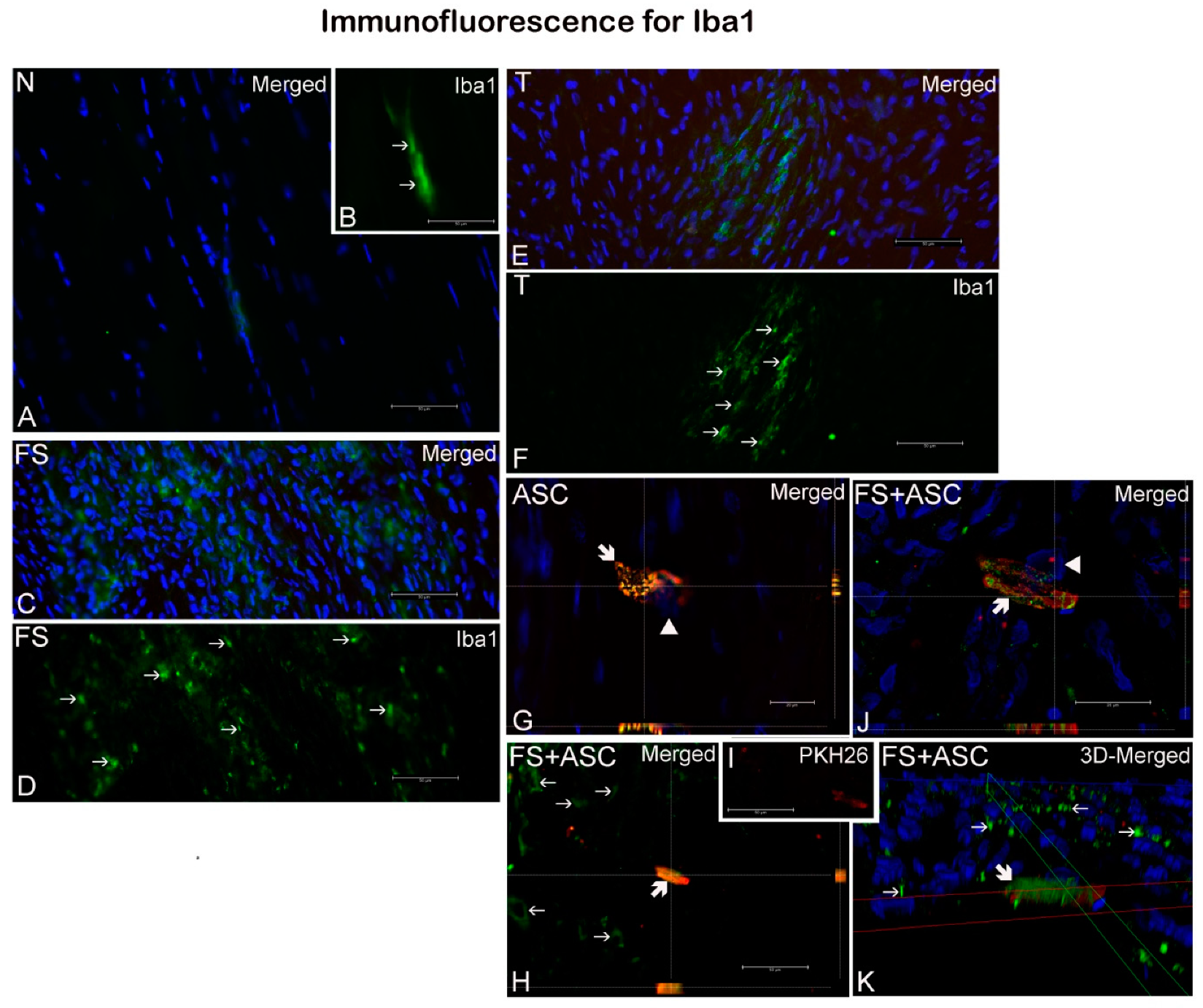
 ). (B) Detail of A, showing Iba1 labeled macrophages. (I) Detail of H, evidencing PHK26 labeled ASC. (G, H, J) orthogonal sectioning; K, 3D deconvolution. Nuclei were stained with DAPI (blue, ▶). Scale bars: A–F, H, I, 50μm; G, J, 20μm.
). (B) Detail of A, showing Iba1 labeled macrophages. (I) Detail of H, evidencing PHK26 labeled ASC. (G, H, J) orthogonal sectioning; K, 3D deconvolution. Nuclei were stained with DAPI (blue, ▶). Scale bars: A–F, H, I, 50μm; G, J, 20μm.
). (B) Detail of A, showing Iba1 labeled macrophages. (I) Detail of H, evidencing PHK26 labeled ASC. (G, H, J) orthogonal sectioning; K, 3D deconvolution. Nuclei were stained with DAPI (blue, ▶). Scale bars: A–F, H, I, 50μm; G, J, 20μm.
). (B) Detail of A, showing Iba1 labeled macrophages. (I) Detail of H, evidencing PHK26 labeled ASC. (G, H, J) orthogonal sectioning; K, 3D deconvolution. Nuclei were stained with DAPI (blue, ▶). Scale bars: A–F, H, I, 50μm; G, J, 20μm.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | N | T | FS | ASC | FS+ASC |
|---|---|---|---|---|---|
| Hydroxyproline (mg/g tissue) | 100.8 ± 7.5 * | 103.6 ± 9.1 | 104.9 ± 8.5 | 107.9 ± 6.5 * | 101.6 ± 6.7 |
| Groups | N | T | FS | ASC | FS + ASC |
|---|---|---|---|---|---|
| Fibroblasts | 26.6 ± 3.8 * | 88.7 ± 11.7 * | 93.9 ± 9.8 * | 96.7 ± 12.3 * | 90.5 ± 14.2 * |
| N | T | FS | ASC | FS + ASC | |
|---|---|---|---|---|---|
| In vivo imaging for ASC detection on tendon | - | - | - | Detection of labeled-ASC until the 7th day | Detection of labeled-ASC until the 14th day |
| Cell migration assay | - | - | - | Presence of ASC on the 21st day | Presence of ASC on the 21st day |
| Macrophages identification | Presence of few macrophages | Presence of few macrophages | Presence of few macrophages | Presence of few macrophages carring PKH26 signal | Presence of few macrophages carring PKH26 signal |
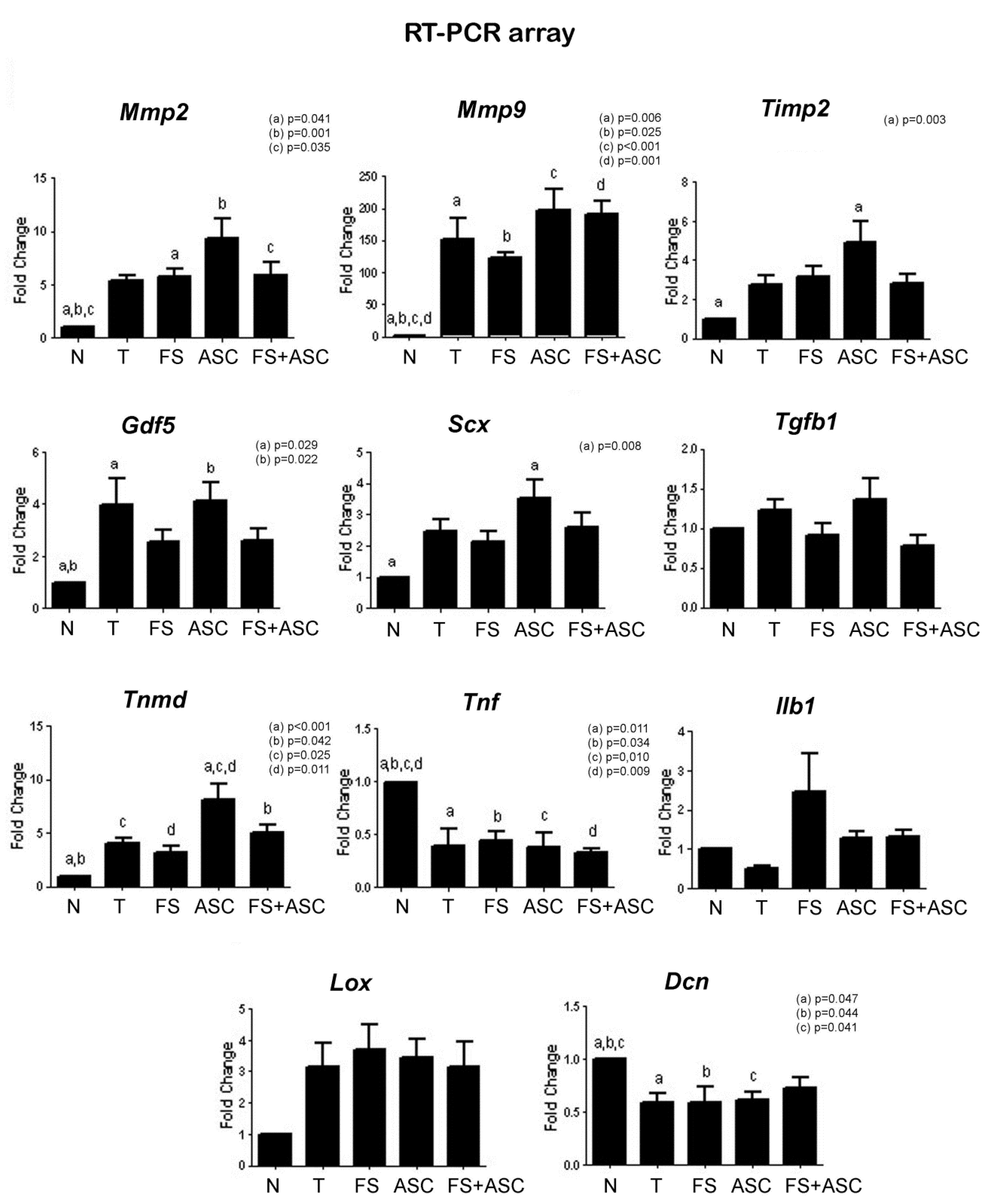
| Genes expression analysis | Lower expression of Mmp2, Mmp9, Timp2, Gdf5, Scx and Tnmd; and higher expression of Tnf and Dcn in relation to T, FS, ASC and FS + ASC groups | Lower Tnmd expression in relation to ASC group | Lower Tnmd expression in relation to ASC group | Higher Tnmd expression in relation to T and FS groups | No differences between the treatments |
| Total collagen concentration (entire tendon) | Lower concentration in relation to ASC group | No differences between the treatments and N group | No differences between the treatments and N group | No differences between the treatments and higher concentration in relation to N group | No differences between the treatments and N group |
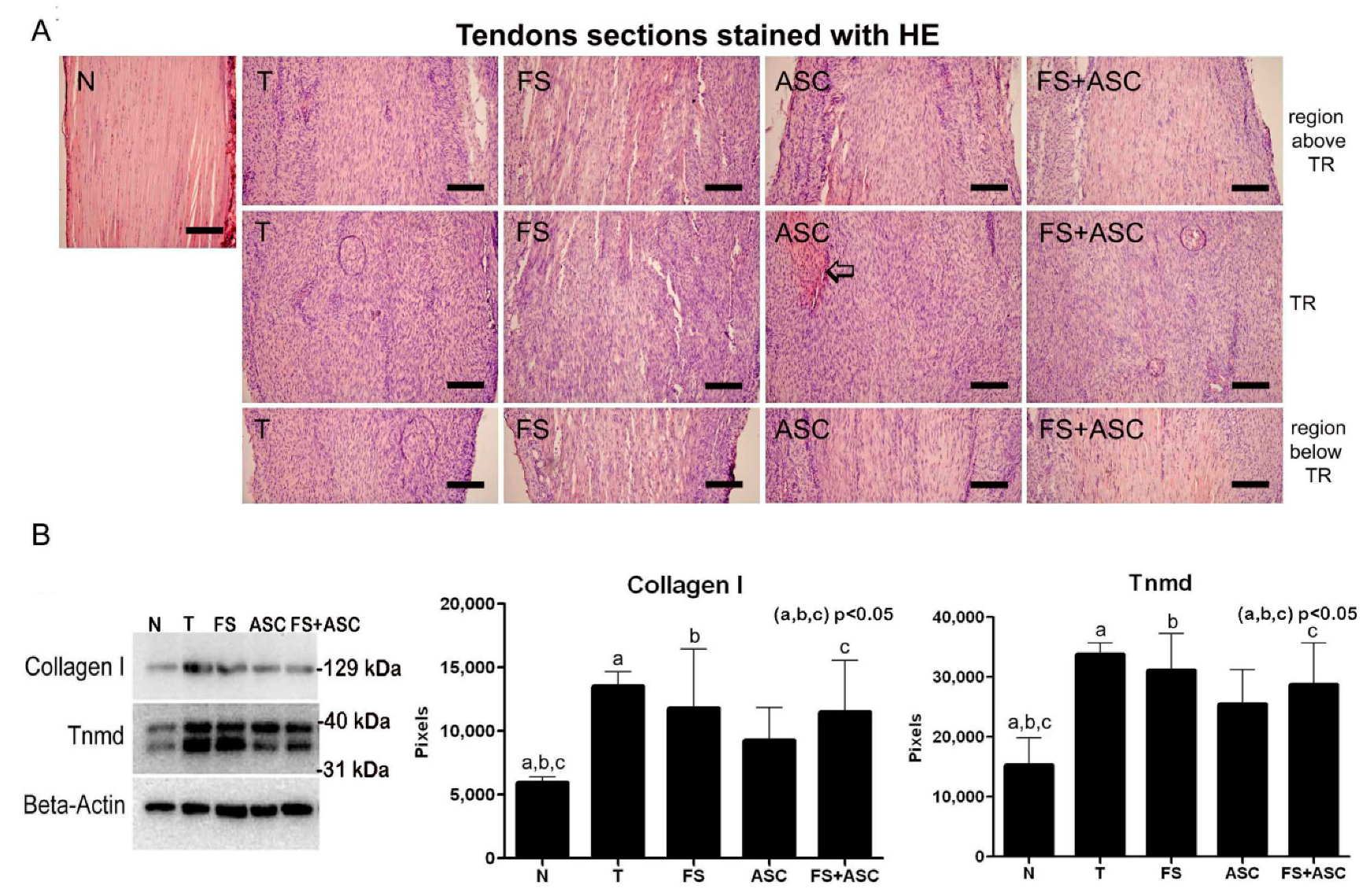
| Collagen I and Tnmd quantification (entire tendon) | Lower amount in relation to T, FS and FS+ASC groups | No differences between the treatments | No differences between the treatments | No differences between the treatments and N group | No differences between the treatments |
| Total number of fibroblasts in the TR | Higher number of cells in relation to T, FS, ASC and FS+ASC groups | No differences between the treatments | No differences between the treatments | No differences between the treatments | No differences between the treatments |
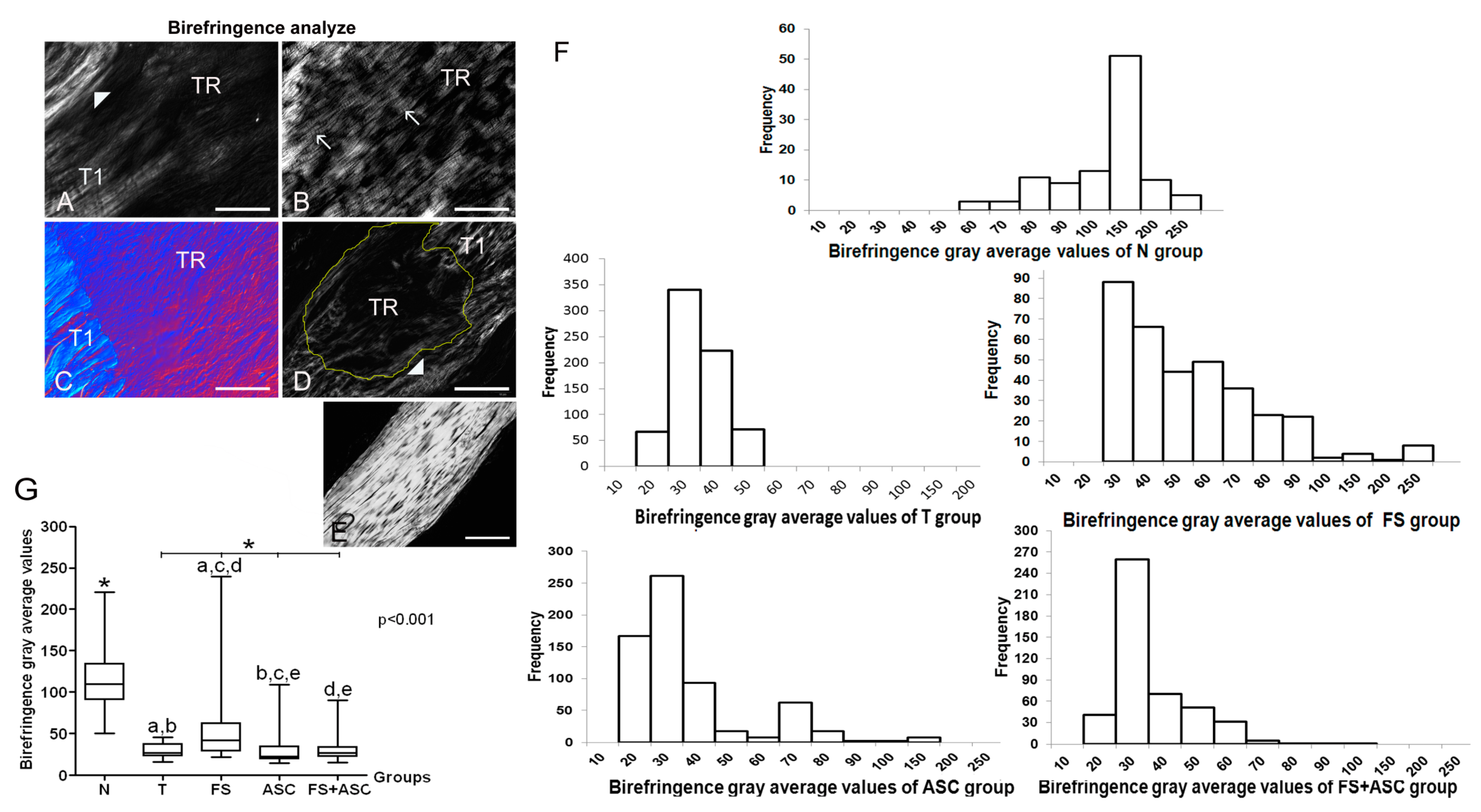
| Collagen fibers organization measurements | Higher birefringence in relation to T, FS, ASC and FS+ASC groups | Lower birefringence in relation to FS, ASC and FS+ASC groups | Higher birefringence in relation to T, ASC and FS+ASC groups | Higher birefringence in relation to T group | Higher birefringence in relation to T and ASC groups |
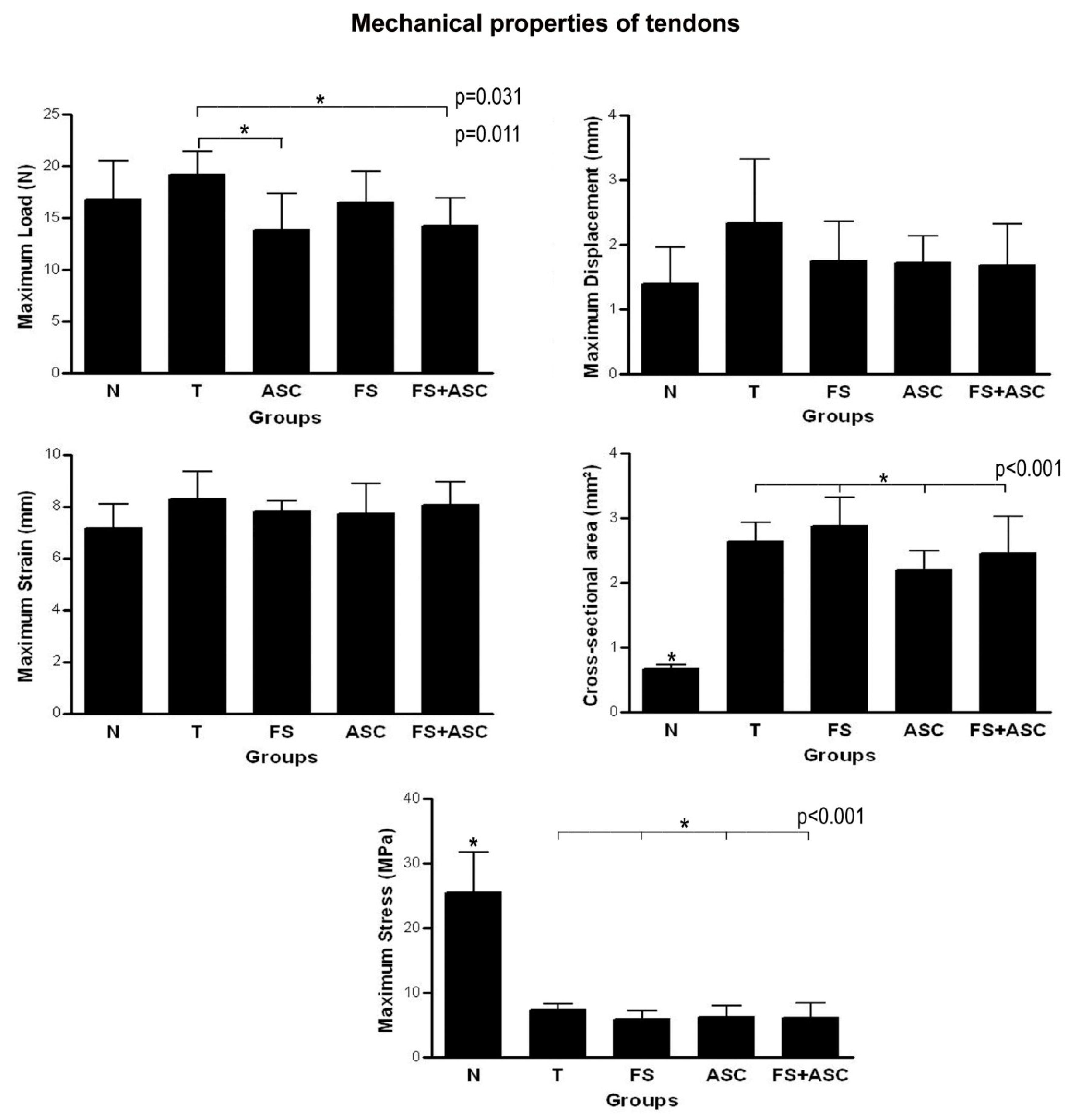
| Biomechanical properties of tendons | Maximum Load: no differences in relation to T, FS, ASC and FS+ASC groups; Cross-sectional area: lower value in relation to T, FS, ASC and FS+ASC groups; Maximum Displacement and Strain: no differences in relation to T, FS, ASC and FS+ASC groups; Maximum Stress: higher value in relation to T, FS, ASC and FS+ASC groups | Maximum Load: higher value in relation to ASC and FS+ASC groups; Cross-sectional area: no differences between the treatments; Maximum Displacement and Strain: no differences between the treatments; Maximum Stress: no differences between the treatments | Maximum Load: no differences between the treatments; Cross-sectional area: no differences between the treatments; Maximum Displacement and Strain: no differences between the treatments; Maximum Stress: no differences between the treatments | Maximum Load: lower value in relation to T group; Cross-sectional area: no differences between the treatments; Maximum Displacement and Strain: no differences between the treatments; Maximum Stress: no differences between the treatments | Maximum Load: lower value in relation to T group; Cross-sectional area: no differences between the treatments; Maximum Displacement and Strain: no differences between the treatments; Maximum Stress: no differences between the treatments |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frauz, K.; Teodoro, L.F.R.; Carneiro, G.D.; Cristina da Veiga, F.; Lopes Ferrucci, D.; Luis Bombeiro, A.; Waleska Simões, P.; Elvira Alvares, L.; Leite R. de Oliveira, A.; Pontes Vicente, C.; et al. Transected Tendon Treated with a New Fibrin Sealant Alone or Associated with Adipose-Derived Stem Cells. Cells 2019, 8, 56. https://doi.org/10.3390/cells8010056
Frauz K, Teodoro LFR, Carneiro GD, Cristina da Veiga F, Lopes Ferrucci D, Luis Bombeiro A, Waleska Simões P, Elvira Alvares L, Leite R. de Oliveira A, Pontes Vicente C, et al. Transected Tendon Treated with a New Fibrin Sealant Alone or Associated with Adipose-Derived Stem Cells. Cells. 2019; 8(1):56. https://doi.org/10.3390/cells8010056
Chicago/Turabian StyleFrauz, Katleen, Luis Felipe R. Teodoro, Giane Daniela Carneiro, Fernanda Cristina da Veiga, Danilo Lopes Ferrucci, André Luis Bombeiro, Priscyla Waleska Simões, Lúcia Elvira Alvares, Alexandre Leite R. de Oliveira, Cristina Pontes Vicente, and et al. 2019. "Transected Tendon Treated with a New Fibrin Sealant Alone or Associated with Adipose-Derived Stem Cells" Cells 8, no. 1: 56. https://doi.org/10.3390/cells8010056