Divergence, Convergence and Phenotypic Diversity of Neotropical Frugivorous Bats


Grupo de Investigación en Ecología Animal, Departamento de Biología, Universidad del Valle, Calle 13 100-00, AA 25360, Cali 760001, Colombia
*
Author to whom correspondence should be addressed.
Diversity 2018, 10(3), 100; https://doi.org/10.3390/d10030100
Submission received: 23 May 2018
/
Revised: 28 August 2018
/
Accepted: 29 August 2018
/
Published: 10 September 2018
(This article belongs to the Special Issue Diversity and Conservation of Bats)
Abstract
:Knowing how adaptation shapes morphological evolution is fundamental to understanding the processes that promote biological diversity. However, there is a lack of empirical evidence on the effects of adaptive radiations on phenotypic diversity, which is related to processes that promote phenotypic divergence and convergence. We applied comparative methods to identify shifts in adaptive peaks and to detect divergence and convergence in skull morphology of frugivorous bats (Phyllostomidae: Stenodermatinae and Carollinae), an ecologically diverse group with strong association between skull morphology, feeding performance and diet that suggests adaptive diversification through morphological innovation. We found divergence and convergence for skull morphology. Fifteen peak shifts were found for jaws, which result in four convergent and four divergent regimes. For skull, nine peak shifts were detected that result in three convergent and three divergent regimes. Furthermore, convergence was significant and strong for skull morphology since distantly related organisms converged to the same adaptive optima. Results suggest that convergence indicates the effect of restriction on phenotypes to keep the advantages provided by the skull phenotype that played a central role in the evolution of strict frugivory in phyllostomids. We conclude that convergence has limited phenotypic diversity of functional traits related to feeding in phyllostomid frugivores.
1. Introduction
Understanding how morphological evolution is shaped by adaptation, in the context of phylogenetic history [1,2,3], is important to understanding the processes that promote biological diversity. In particular, the evolution of ecological diversity within a rapidly multiplying linage, or adaptive radiation, is vital to species diversification since it includes the origin of new species and the evolution of ecological differences between them [2,4,5]. Adaptive radiations illustrate the mechanisms that relate adaptation and speciation [2,6], and can be triggered by the ecological opportunity of gaining access to resources free of competitors [6,7,8]. Ecological opportunities can arise by colonization of a new environment with a species-poor biota (i.e., islands or lakes), by extinction of antagonists or by acquisition of a key innovation [9]; and is considered the primary environmental driver of phenotypic evolution and adaptive diversification of species [2,10,11,12]. Nevertheless, an outstanding gap in the study of adaptive radiations is the contribution of ecological opportunity to morphological diversity, which has been poorly quantified even though phenotypic divergence is an important part of the original ecological opportunity hypothesis [2]. Consequently, there is a lack of empirical evidence regarding the influence of ecological opportunities driven by key innovations on phenotypic diversity.
Phenotypic diversification of clades is related to processes that promote morphological divergence and convergence. Phylogenetic conservatism can constrain ecological and morphological diversity inside clades by retaining environmental niches and conserving morphological patterns of a shared inherited lifestyle [13]. On the other hand, adaptation to particular environments might be able to override phylogenetic conservatism [14] Thus, different selective pressures imposed by the environment can generate divergence within a clade by driving phenotypic evolution towards different optima [15], whereas similar selective pressures can lead to convergence among distantly related species [16]. Convergent phenotypes are common between species from different lineages that share a similar ecotype [17,18,19,20]. The study of convergent evolution is essential to understand how natural selection optimizes an organism for a specific niche [21]. In particular, the study of species-rich groups can improve the ability to clearly identify repeated patterns of morphological evolution and the processes involved.
Phyllostomid diversity has originated through a possible mechanism of adaptive ecological diversification through morphological innovation [4], which is supported by the strong associations between skull morphology, bite force and diet in these bats [22,23,24]. Functional demands imposed by dietary specializations have influenced cranial evolution during the radiation of phyllostomid bats [4,25,26,27], suggesting strong clade-based links between ecological opportunity and diversification [4]. In particular, Stenodermatinae represents a different adaptive zone by having a diet that contains significantly more fruit than that of other phyllostomids, and has a markedly higher diversification rate than the others subfamilies [4,27]. Strict frugivory, in Stenodermatinae, is related to consumption of soft and hard fruits [4], which is allowed by a strong bite that is, in turn, associated to a short and wide skulls [4,24,26]. Fruit feeders are morphologically diverse, exhibiting cranial and mandibular morphologies that overlap with other guilds [25]. This morphological diversity reflects the variety of physical properties represented among fruits, and the tendency of frugivorous species to specialize on particular fruits [24,25,28]. However, the role of adaptive radiations on ecological and morphological diversification of frugivorous bats and the extent to which functional traits related to feeding reflect convergence or divergence is poorly known (but see Santana et al., 2010 [24] and Rossoni et al. [28]).
Consequently, to better understand the role of adaptive radiations on morphological diversification, we assessed the extent to which the skull size and shape of frugivorous phyllostomid bats has been driven by morphological convergence and divergence. Specifically, we characterized divergence and convergence in skull morphology (size and shape) of phyllostomid frugivores and tested for the strength of convergent evolution in functional traits related to fruit feeding. Our analyses support a central role of convergence in both keeping the adaptive skull phenotype of Stenodermatinae, which allowed evolution of strict frugivory, and restricting skull diversity within phyllostomid frugivores.
2. Materials and Methods
2.1. Cranial Morphology
We measured 647 individuals of 46 species, which covered 12 of the 18 genera in the Stenodermatinae subfamily, and all the genera in the Carollinae subfamily (Carollia and Rhinophylla) (Table S1). To account for sexual dimorphism, we measured 10 males and females of each species whenever possible. Otherwise, we measured all available specimens. However, we were only able to measure one specimen for Ectophylla alba and Sphaeronycteris toxophyllum (Table S1). All specimens were measured from mammal’s collection of Universidad del Valle, except Centurio senex specimens were measured from mammal’s collection of Universidad Autómona de Mexico (México D.F.). We measured 20 traits of functional units related to feeding (cranium and mandible), which reflect skull shape and size and are associated with feeding performance and diet in phyllostomid bats [4,24,26]: Total skull length (TL), Postorbital Width (PW), Zygomatic Breadth (ZB), Posterior Skull width (PSW), Palatal Width at canines (PW), Palatal Width at first molar (PW1), Total Palate Length (TPL), Anterior Skull Length (ASL), Post-Palatal length (PPL), Maxillary Toothrow Length (MTL), Dentary Depth under first molar (DD), Coronoid Process Height (CPH), Condyle height (CH), Condyle to Canine bite point distance (CC), Condyle to first molar point distance (Cm1), Condyle to last molar point distance (Cm3), Total Dentary Length (TDL), Condyle-Coronoid Length (CCL), Coronoid-Angular Length (CAL) and Mandibular toothrow length (MAN). All morphological measurements were carried out using a digital caliper (precision 0.01 mm). We followed taxonomic designations of species by Gardner [29].
2.2. Convergent Evolution in Neotropical Frugivorous Bats
To quantify cranial morphology, we performed phylogenetic principal components analyses based on the species averages of morphometric cranial traits (separately for skull and jaw) and on the covariance matrix. Then, we used species scores from the first five pPCs to describe cranial size and shape in subsequent analyses. For phylogenetic analyses, we used the phylogeny of Phyllostomidae corresponding to the maximum clade credibility tree used in Rolland et al. [30], which is implemented in RPANDA [31] R-package [32]. This time-calibrated tree includes 150 extant species of Phyllostomidae bats. To perform our phylogenetic comparative analyses, we trimmed the tree to only include the 46 frugivorous bats from Stemodermatinae and Carollinae species we were able to measure (Table S1).
To determine the optimal number of selective regimes on the phylogenetic tree, we applied a model-based approach that uses LASSO (Least Absolute Shrinkage and Selector Operator) and is implemented in the R-package l1ou [33]. This method detects shifts in phenotypic optima on trees using the Ornstein–Uhlenbeck model (OU) [34]. The standard OU process includes random perturbations and stabilizing selection and assumes that species evolve independently. The OU model describe the evolution of a continuous phenotypic trait X(t) over time t by including: (1) a deterministic component that models the tendency of a trait to evolve toward an adaptive optimum (θ(t)), which may change with time and across branches of the phylogeny; and (2) a stochastic component (dB(t)) that models random evolution with linearly increasing variance over time under a Brownian motion (BM) model. Ornstein–Uhlenbeck (OU) process is defined by the following stochastic equation:
where B(t) is the Brownian motion model (BM), in which random deviations are introduced by the Gaussian white noise dB(t) with the magnitude of these deviations depending on the noise intensity parameter σ. In addition, OU model considers trait adaptation to the environment through the optimum trait values (θ(t)) and with the tendency of a trait to evolve toward that adaptive optimum considered by the parameter α, which quantifies the rate at which the trait is pulled toward an optimum and is interpreted as strength of selection. l1ou uses the OU model to quantify the number of selection regimes needed to explain the data, and detects convergent adaptive peaks among distantly related taxa by collapsing similar sets of Ornstein–Uhlenbeck parameters in the same evolutionary regime by using a phylogenetic Bayesian Information Criterion (pBIC) [33]. This is more conservative than those used by other methods such as SURFACE [35]. Thus, this method does not require pre-specification of selective regimes on the phylogeny. We applied l1ou to pPC scores to detect convergence in skull morphology across the phylogeny of frugivorous phyllostomid bats.
2.3. Test and Quantification of Convergence
We applied the Wheatsheaf index (w) [36] to investigate the strength of morphological convergence among the groups of convergent species previously identified with l1ou method. This index compares the mean distance in phenotypic space between all pairs of species with the mean pairwise comparisons between convergent species only, incorporating phylogenetic relatedness [36]:
where is the mean for pairwise comparisons between all species, and is the mean for pairwise comparisons between focal species only. As increases and decreases, w increases, showing stronger convergence and vice versa. We tested whether convergence strength (quantified by the Wheatsheaf index) among species of convergent regimes was significantly stronger than expectations from random distributions of trait values (pPC scores) across the phylogeny by using the windex package [37]. Windex performs a bootstrapping approach to resample the tips of the tree along with their trait values and thus obtain a distribution of possible w indices given the phylogeny and the trait values for each species. Using this distribution and the calculated value of w, we could generate a “p-value” by taking the proportion of bootstrap samples that are greater than or equal to the value of w calculated from the original dataset. Then, this test represents an evaluation of whether convergence is significantly stronger than would expected compared to a random distribution of trait values across the specified phylogeny [37].
3. Results
3.1. Cranial Morphology
The first five phylogenetic principal components (pPCs) explained 99.91% of the variation in jaw morphology of bat species. Factor loads indicated that pPC1 represented jaw size (factor loads were similar and negatives), pPC2 represented dentary depth (MDD = 0.519), pPC3 represented relationship between condyle height (CH = 0.102) and mandibular toothrow length (MAN = −0.129), pPC4 represented condyle to last molar distance (Cm3 = −0.142) and pPC5 represented relations between dentary depth under first molar (MDD = −0.066) and coronoid process height (CPH = 0.071) (Table 1). For cranium, the first five pPCs contained 99.83% of variation in shape. pPC1 represented cranium size (TSL = −0.980, MSW = −0.904, TPL = −0.986, ASL = −0.994, and MTL = −0.995), pPC2 represented relationship between face width (PW = −0.988, PW1 = −0.981, and ZB = −0.718) and post-palatal length (PPL = 0.772), pPC3 represented post-palatal length (PPL = −0.577), pPC4 represented relationship between palatal width (PW1 = −0.153) and post-palatal length (PPL = 0.106), and pPC5 represented relationship between zygomatic breadth (MZB = −0.106) and post-palatal length (PPL = 0.134). Consequently, pPCs were associated with traits that are related to bite force in phyllostomids (skull length and height, condyle height, condyle to first molar distance, zygomatic breadth, posterior skull width, and palate width at first molar), so we used the first five pPCs to represent cranial attributes related to bite performance in subsequent analysis.
3.2. Convergent Evolution in Neotropical Frugivorous Bats
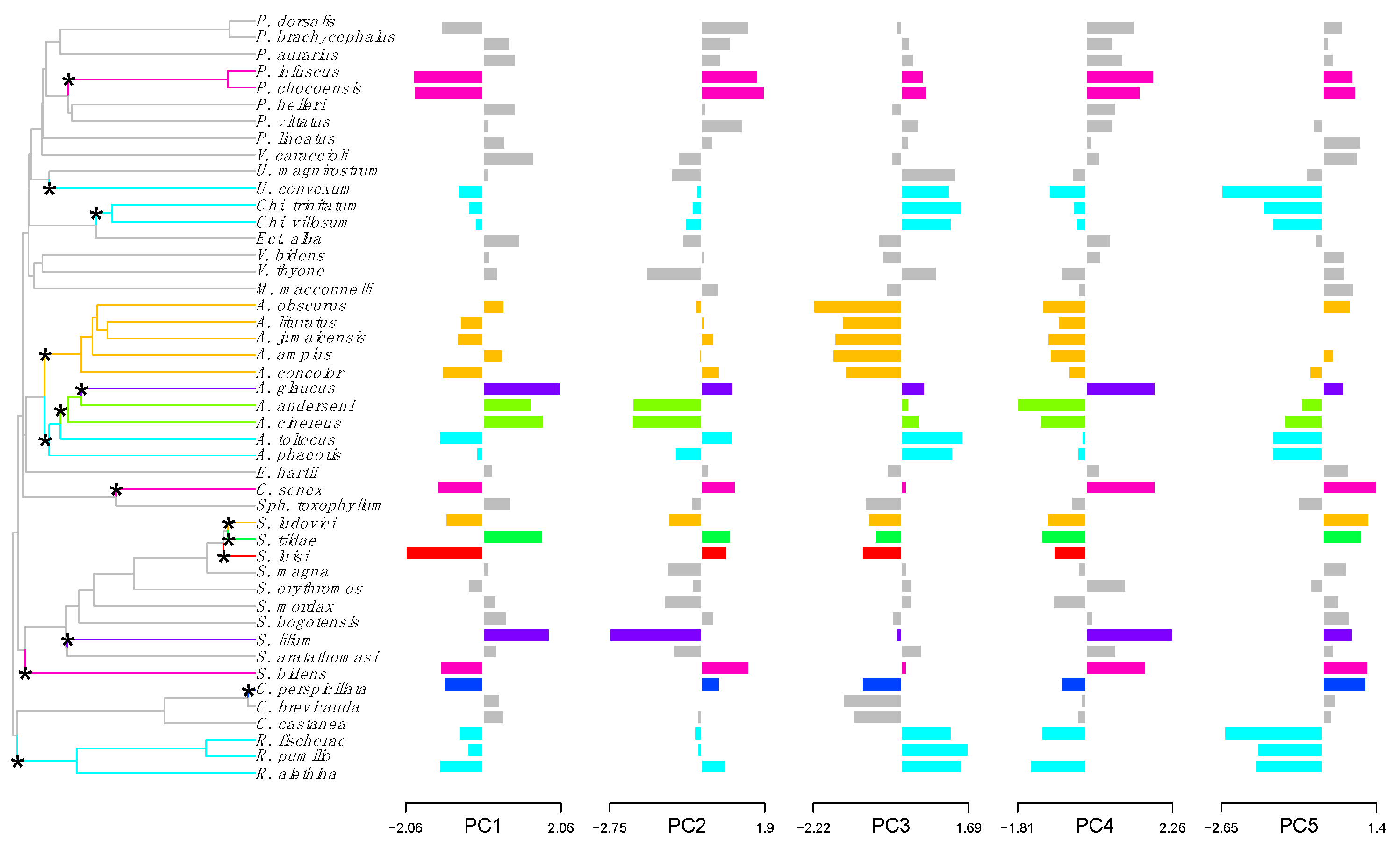
According to l1ou analyses, adaptive shifts in skull and jaw morphology are common across the phylogeny of phyllostomid frugivorous bats (subfamilies Stenodermatinae and Carollinae). These shifts correspond to multivariate combinations of the traits used for the analyses (pPCs); thus, each of these shifts is supported by several of the five traits used (Figure 1 and Figure 2). For jaw, 15 adaptive shifts in shape were detected (most with bootstrap support values >70%, Figure 1) and a total of eight evolutionary regimes were estimated across the phylogeny since 11 shifts result in four convergent regimes for jaw (Figure 1): (1) Platyrrhinus infuscus-P. chocoensis (93.6%), Centurio senex (79.4%) and Sturnira bidens (54.2%) have enlarged jaw, thick dentary, a short condyle to last molar distances and low coronoid process; (2) Uroderma convexum (96.2%), Chiroderma trinitatum-Ch. villosum (89.6%), Artibeus toltecus-A. phaeothis (77.2%), and Rhinophylla fisherae-R. pumilio-R. alethina (99.7%) have high condyle, short mandibular toothrow and high coronoid process; (3) Artibeus obscurus-A. lituratus-A. jamaicenscis-A. amplus-A. concolor (89.7%), and Sturnira ludovici (49.1%) have low condyle, long mandibular toothrow and long condyle to last molar distance; and (4) Artibeus glaucus (98.1%) and Sturnira lilium (94.9%) are small species with small condyle to last molar distance. One of these convergent regimes (2) includes genera from both subfamilies: Artibeus, Chiroderma and Uroderma from Stenodermatinae and Rhinophylla from Carollinae.
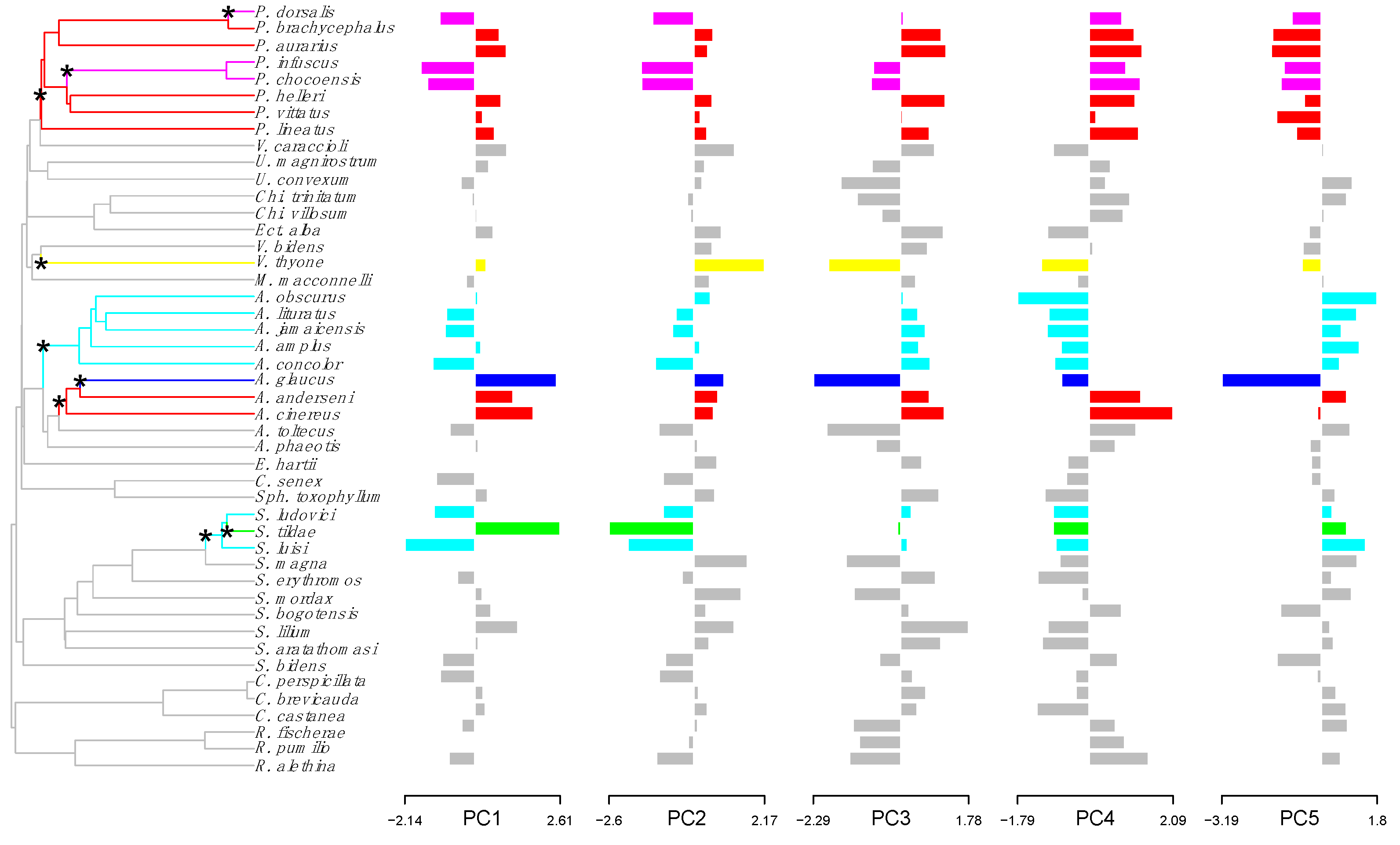
For skull, nine adaptive peak shifts were detected (eight with bootstrap support values >70%, Figure 2) and six regimes were detected since six adaptive shifts result in three convergent regimes for skull (Figure 2): (1) Platyrrhinus dorsalis (51.8%) and P. infuscus-P. chocoensis (71.1%) have long and wide cranium with long a palate; (2) Platyrrhinus brachycephalus, P. aurarius, P. helleri-P. vittatus, P. lineatus (76.3%), and Artibeus anderseni-A. cinereus (75.1%) have short cranium with a short and wide rostrum; and (3) Artibeus obscurus-A. lituratus-A. jamaicencis-A. amplus-A. concolor (76.2%), and Sturnira ludovici-S. luisi have a short cranium with a wide palate (79.5%). All convergent regimes for skull involved only genera of Stenodermatinae.
Fourteen species, from the three most diverse genera (Artibeus, Platyrrhinus and Sturnira), showed shifts in adaptive peaks for both jaw and skull (Figure 1 and Figure 2): A. anderseni, A. cinereus, A. obscurus, A. lituratus, A. jamaicenscis, A. amplus, A. concolor; A. glaucus, P. infuscus, P. chocoensis, S. ludovici, S. luisi and S. tildae.
3.3. Test and Quantification of Convergence
The Wheatsheaf index indicated convergence for cranial size and shape in adaptive shifts, identified by l1ou method, was significantly stronger than expected compared to a random distribution (Table 2 and Table 3). In addition, convergence strength was higher for jaw than for skull since values of the Wheatsheaf index were higher and more variables were significant (p < 0.05) for jaw than for skull (Table 2 and Table 3). However, the third convergent regime of jaw (Artibeus obscurus-A. lituratus-A. jamaicenscis-A. amplus-A. concolor) was not supported by data since Wheatsheaf index value was not different than expected by chance (Table 2 and Table 3).
4. Discussion
We found several shifts in adaptive peaks across the phylogeny of Neotropical frugivorous bats (Phyllostomidae: Stenodermatinae and Carollinae), which indicate divergence in cranium and jaw morphology. In particular, the most diverse genera showed a higher level of intrageneric divergence in morphology than less diverse genera. On the other hand, the degree of convergence was stronger for jaw than for cranium. Therefore, our results suggest that convergence can result of maintaining the skull shape that drove diversification in Stenodermatinae, which may have limited the variation in skull phenotypes by driving distantly related organisms towards similar adaptive optima.
The presence of shifts in adaptive peaks inside all clades indicated significant divergence in cranial morphology; this divergence was higher for jaws (12 adaptive shifts) than for skulls (8 adaptive shifts). Additionally, several species showed simultaneous occurrence of adaptive shifts for jaw and skull, which suggests cranial integration of functional traits related to feeding. Jaw and skull morphology is related with food loading and processing because of muscle insertion and teeth support [38,39,40,41]. In particular, Stenodermatinae bats showed divergent morphologies inside clades, which was evident for species-rich genera such as Platyrrhinus, Sturnira and Artibeus. On the other hand, less-diverse Carollinae genera, Carollia and Rhinophylla, exhibited less divergence in cranial phenotype. Thus, results suggest that divergence between closely related species, which can increase the set of adaptations for consumption of different fruit types, could promote diversification by generating divergent phenotypes within a lineage. Stenodermatinae clade has shown high rates of speciation, which has been associated to a short and wide cranium that confers increase in bite force [4,24,26], allowing the consumption of soft and hard fruits and the benefit from the ecological opportunity of strict frugivory [4]. For example, Artibeus jamaicensis eats large quantities of hard fruits such as figs when these are available (seasonally) and consumes soft fruits throughout the year [42], so this species depends largely on a frugivore diet. Morphological traits associated with divergence in this study (zygomatic amplitude, palatal amplitude and the post palatal longitude) have been identified as important for developing the frugivore diet of Stenodermatinae bats [4]. Therefore, results suggest that variation in cranial and mandible morphology of fruit bats can be the results of adaptive pressures related to requirements for consumption of fruits with diverse physical properties. Even though we have not presented evidence that the adaptive peaks found are associated with diet in bats, studies have supported the hypothesis that most of the variation in phyllostomid skull form and biomechanics is underlined by broad changes in dietary hardness [26]. Consequently, the presence of multiple adaptive peaks of cranial phenotypes in Stenodermatinae bats supports the hypothesis of an adaptive radiation driven by diet specialization (i.e., [4,27]). This hypothesis proposes that acquisition of a skull phenotype that improved biting performance in Stenodermatinae, opened a new adaptive zone that increased diversification rates in these bats [4,26]. Furthermore, adaptive shifts of species from different clades of Stenodermatinae converged to the same phenotypic adaptive optima; this convergence was strong for both jaw and skull.
Mapping morphology on the phylogeny of frugivorous bats revealed that similar phenotypes have evolved repeatedly across clades; this indicates morphological convergence and suggests that many fruit specialist replicates have been evolved in Stenodermatinae. Additionally, the degree of convergence between the multiple adaptive peaks that were detected was different than expected based on null models for both skull and jaw. Evolutionary convergence results from exposure to similar selective pressures, which favor morphological and physiological traits that maximize fitness and competitive ability under those pressures [21]. Results showed that convergent regimes were associated with traits related to bite force such as jaw and dentary length and height of mandibular condyles in the mandible, and with cranium length and width, which suggest that there is convergence for similar bite performance and that these phenotypes may be adaptive and represent true convergence. The fact that bats show similar cranial and mandible shape indicates that they can have similar bite performance, which can result in consumption of fruit with similar physical properties. On the other hand, bat species differ in many ecological traits (such as body size, echolocation calls, olfactive capability, vision acuity, flight maneuverability, etc.) that can influence foraging strategies and so fruit selection. Consequently, even if they show convergence in morphology, they not necessarily must have similar taxonomic diet, but we expect that they consume fruits with similar physical properties (especially if they have similar body size). The indication that selection has acted on skull function and has caused convergence on morphology opens the door for future studies that investigate how similar is diet among morphologically convergent species. However, to identify functional dietary groups of phyllostomid bats that at least grossly match morphological differences of their skull, more quantitative data on the hardness of food items consumed by all animals is needed. Therefore, based on the strong association among diet, skull morphology and feeding performance [4,24,25,26,43,44], our results suggest that divergence inside clades leads to convergence in skull shape between species of different clades, which can indicate phenotypes with similar biting performance that can eventually conduct to consumption of fruit with similar physical properties.
Parallel adaptive evolution of species that are part of the same broader radiation is relatively common [45], and it often leads to morphological convergence due to independent adaptations to the same adaptive niche [2,21,46]. Stenodermatinae cranial phenotype is a novel trait that has played a central role in the speciation within phyllostomids (highly taxonomically and ecologically diverse mammal family) by promoting the evolution of strict frugivory [4]. Thus, our results suggest that constraints to maintain the key innovation, that opened the ecological opportunity of strict frugivory, could drive distantly related fruit bats towards the same phenotypic adaptive optima; which agree with previous results that cranial morphology has evolved more slowly in Stenodermatinae than in other phyllostomids [4]. Adaptive radiations increase morphological diversity inside clades, but convergence among them restricts the overall morphological variation as suggests results from frugivorous Phyllostomids. This convergence can limit the suite of phenotypic traits that will evolve as adaptations. Consequently, divergence inside fruit bat clades promotes intra-clade diversity but restrictions due to the maintaining of the Stenodermatinae skull phenotype results in convergence in shape with species from other linages. Bats are among the most diverse local mammals in tropical communities, with Phyllostomidae being the dominant family, consequently phyllostomids can be subject to strong competition for resources [47]. Thus, resource competition and ecological character displacement can potentially play a major role in morphological, ecological and evolutionary diversification by promoting morphological specialization. However, evolution of different cranial morphologies along with differences in sensory capabilities, behavior and body sizes can expand their niche and experience a reduction in interspecific competition among species of the same clade, which can allow coexistence without morphological consequences. In addition, seasonal variation in food availability and competition with bats might maintaining selection on morphological traits that grant access to hard foods. Therefore, understanding convergent evolution is important to understanding the generation of biodiversity, constraints on adaptation, and how natural selection optimizes an organism for a particular niche.
Ecological opportunities have influenced speciation rate of contemporary linages [4,5,10,12,48], but the relationship between ecological opportunity and phenotypic divergence has been poorly quantified in contemporary taxa [2]. We found diverse shifts in adaptive peaks across the phylogeny of Neotropical frugivorous bats (Phyllostomidae: Stenodermatinae and Carollinae), which indicate divergence for skull and jaw morphology. In particular, species of the most diverse genera showed a significant level of intrageneric divergence in morphological with respect to that of less diverse genera. Therefore, results suggest that variation in cranial and mandible morphology of fruit bats can be the results of adaptive pressures related to requirements for consumption of different types of fruits. Furthermore, we found divergent selection inside clades of frugivorous bats that led to evolution of convergent cranial phenotypes. The fact that we found similar skull morphology in distantly related Stenodermatinae bats indicates the effect of restriction on phenotypes to keep the advantages provided by the cranial phenotype that allow strict frugivory in Phyllostomidae. However, even though new morphotypes can appear in a particular clade [2], this evolution of different phenotypes does not provide new ecological niches for the overall linage. Consequently, results suggest that convergence between organisms for particular niches can limit the variation in phenotypes by driving distantly related organisms towards the same phenotypic adaptive optima.
Supplementary Materials
The following are available online at https://www.mdpi.com/1424-2818/10/3/100/s1, Table S1: Bats species used in the analyses, and Phylogenetic Principal Components (pPC) scores for jaw and cranium.
Author Contributions
O.E.M-G. conceived the idea, analyzed data and wrote the manuscript. M.E.D.l.v. measured the specimens and wrote the manuscript.
Funding
This research received no external funding.
Acknowledgments
We thank Luisa A. Ruano for measuring specimens of Centurio senex. We also thank Fernando A. Cervantes for allowing the measurement of specimens from Colección Nacional de Mamíferos of Universidad Autómona de Mexico (México D.F.)
Conflicts of Interest
The authors declare no conflict of interest.
References
- Losos, J.B. The evolution of form and function: Morphology and locomotor performance in West Indian Anolis lizards. Evolution 1990, 44, 1189–1203. [Google Scholar] [CrossRef] [PubMed]
- Schluter, D. The Ecology of Adaptive Radiation; Oxford University Press: New York, NY, USA, 2000; ISBN 9780191588327. [Google Scholar]
- Gavrilets, S.; Losos, J.B. Adaptive radiation: Contrasting theory with data. Science 2009, 323, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Dumont, E.R.; Davalos, L.M.; Goldberg, A.; Santana, S.E.; Rex, K.; Voigt, C.C. Morphological innovation, diversification and invasion of a new adaptive zone. Proc. R. Soc. B Biol. Sci. 2012, 279, 1797–1805. [Google Scholar] [CrossRef] [PubMed]
- Parent, C.E.; Crespi, B.J. Ecological opportunity in adaptive radiation of Galápagos endemic land Snails. Am. Nat. 2009, 174, 898–905. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B. Adaptive radiation, ecological opportunity, and evolutionary determinism. Am. Nat. 2010, 175, 623–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellborn, G.A.; Langerhans, R.B. Ecological opportunity and the adaptive diversification of lineages. Ecol. Evol. 2015, 5, 176–195. [Google Scholar] [CrossRef] [PubMed]
- Burress, E.D.; Holcomb, J.M.; Tan, M.; Armbruster, J.W. Ecological diversification associated with the benthic-to-pelagic transition by North American minnows. J. Evol. Biol. 2017, 30, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Simpson, G.G. The Major Features of Evolution; Columbia University Press: New York, NY, USA, 1953. [Google Scholar]
- Mahler, D.L.; Revell, L.J.; Glor, R.E.; Losos, J.B. Ecological opportunity and the rate of morphological evolution in the diversification of greater Antillean anoles. Evolution 2010, 64, 2731–2745. [Google Scholar] [CrossRef] [PubMed]
- Nosil, P.; Reimchen, T.E. Ecological opportunity and levels of morphological variance within freshwater stickleback populations. Biol. J. Linn. Soc. 2005, 86, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Harmon, L.J.; Melville, J.; Larson, A.; Losos, J.B. The role of geography and ecological opportunity in the diversification of day geckos (Phelsuma). Syst. Biol. 2008, 57, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Wiens, J.J.; Graham, C.H. Niche Conservatism: Integrating evolution, ecology, and conservation biology. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 519–539. [Google Scholar] [CrossRef]
- Moen, D.S.; Irschick, D.J.; Wiens, J.J. Evolutionary conservatism and convergence both lead to striking similarity in ecology, morphology and performance across continents in frogs. Proc. R. Soc. B Biol. Sci. 2013, 280, 20132156. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B.; Warheitt, K.I.; Schoener, T.W. Adaptive differentiation following experimental island colonization in Anolis lizards. Nature 1997, 387, 70–73. [Google Scholar] [CrossRef]
- Rüber, L.; Adams, D.C. Evolutionary convergence of body shape and trophic morphology in cichlids from Lake Tanganyika. J. Evol. Biol. 2001, 14, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, A.Y.; Rubinoff, D. Convergent evolution of morphology and habitat use in the explosive Hawaiian fancy case caterpillar radiation. J. Evol. Biol. 2013, 26, 1763–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingram, T.; Kai, Y. The geography of morphological convergence in the radiations of Pacific Sebastes rockfishes. Am. Nat. 2014, 184, E115–E131. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.T.; Price, S.A.; Hoey, A.S.; Wainwright, P.C. Ecomorphological convergence in planktivorous surgeonfishes. J. Evol. Biol. 2016, 29, 965–978. [Google Scholar] [CrossRef] [PubMed]
- McKown, A.D.; Akamine, M.E.; Sack, L. Trait convergence and diversification arising from a complex evolutionary history in Hawaiian species of Scaevola. Oecologia 2016, 181, 1083–1100. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B. Convergence, adaptation, and constraint. Evolution 2011, 65, 1827–1840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumont, E.R.; Herrel, A.; Medellín, R.A.; Vargas-Contreras, J.A.; Santana, S.E. Built to bite: Cranial design and function in the wrinkle-faced bat. J. Zool. 2009, 279, 329–337. [Google Scholar] [CrossRef]
- Aguirre, L.F.; Herrel, A.; Van Damme, R.; Matthysen, E. The implications of food hardness for diet in bats. Funct. Ecol. 2003, 17, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Santana, S.E.; Dumont, E.R.; Davis, J.L. Mechanics of bite force production and its relationship to diet in bats. Funct. Ecol. 2010, 24, 776–784. [Google Scholar] [CrossRef] [Green Version]
- Dumont, E.R. Cranial shape in fruit, nectar, and exudate feeders: Implications for interpreting the fossil record. Am. J. Phys. Anthropol. 1997, 102, 187–202. [Google Scholar] [CrossRef]
- Santana, S.E.; Grosse, I.R.; Dumont, E.R. Dietary hardness, loading behavior, and the evolution of skull form in bats. Evolution 2012, 66, 2587–2598. [Google Scholar] [CrossRef] [PubMed]
- Rojas, D.; Vale, Á.; Ferrero, V.; Navarro, L. The role of frugivory in the diversification of bats in the Neotropics. J. Biogeogr. 2012, 39, 1948–1960. [Google Scholar] [CrossRef]
- Rossoni, D.M.; Assis, A.P.A.; Giannini, N.P.; Marroig, G. Intense natural selection preceded the invasion of new adaptive zones during the radiation of New World leaf-nosed bats. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gardner, A. Mammals of South. America, Volume 1: Marsupials, Xenarthrans, Shrews, and Bats; The University of Chicago Press: Chicago, IL, USA, 2008; ISBN 9780226282404. [Google Scholar]
- Rolland, J.; Condamine, F.L.; Jiguet, F.; Morlon, H. Faster speciation and reduced extinction in the tropics contribute to the mammalian latitudinal diversity gradient. PLoS Biol. 2014, 12, e1001775. [Google Scholar] [CrossRef] [PubMed]
- Morlon, H.; Lewitus, E.; Condamine, F.L.; Manceau, M.; Clavel, J.; Drury, J. RPANDA: An R package for macroevolutionary analyses on phylogenetic trees. Methods Ecol. Evol. 2016, 7, 589–597. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Hansen, T.F. Stabilizing selection and the comparative analysis of adaptation. Evolution 1997, 51, 1341–1351. [Google Scholar] [CrossRef] [PubMed]
- Khabbazian, M.; Kriebel, R.; Rohe, K.; Ané, C. Fast and accurate detection of evolutionary shifts in Ornstein-Uhlenbeck models. Methods Ecol. Evol. 2016, 7, 811–824. [Google Scholar] [CrossRef]
- Ingram, T.; Mahler, D.L. SURFACE: Detecting convergent evolution from comparative data by fitting Ornstein-Uhlenbeck models with stepwise Akaike Information Criterion. Methods Ecol. Evol. 2013, 4, 416–425. [Google Scholar] [CrossRef]
- Arbuckle, K.; Bennett, C.M.; Speed, M.P. A simple measure of the strength of convergent evolution. Methods Ecol. Evol. 2014, 5, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Arbuckle, K.; Minter, A. Windex: Analyzing convergent evolution using the wheatsheaf index in R. Evol. Bioinforma. 2015, 11, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Herring, S.W.; Rafferty, K.L.; Liu, Z.J.; Marshall, C.D. Jaw muscles and the skull in mammals: The biomechanics of mastication. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 131, 207–219. [Google Scholar] [CrossRef]
- Cox, P.G. A quantitative analysis of the Eutherian orbit: Correlations with masticatory apparatus. Biol. Rev. 2008, 83, 35–69. [Google Scholar] [CrossRef] [PubMed]
- Pitnick, S.; Jones, K.E.; Wilkinson, G.S. Mating system and brain size in bats. Proc. R. Soc. B Biol. Sci. 2006, 273, 719–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellens, H.L.L.; Kuijpers-Jagtman, A.M.; Halazonetis, D.J. Geometric morphometric analysis of craniofacial variation, ontogeny and modularity in a cross-sectional sample of modern humans. J. Anat. 2013, 222, 397–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handley, C.O.; Wilson, D.E.; Gardner, A.L. Demography and Natural History of the Common Fruit Bat, Artibeus Jamaicensis, on Barro Colorado Island, Panamá; Smithsonian Institution Press: Washington, DC, USA, 1991; ISBN 0081-0282. [Google Scholar]
- Dumont, E.R. The correlated evolution of cranial morphology and feeding behavior in New World fruit bats. In Functional and Ecological Morphology of Bats; Kunz, G., McCraken, G., Akbar, Z., Eds.; Oxford University Press: New York, NY, USA, 2006; pp. 160–177. ISBN 139780195154726. [Google Scholar]
- Freeman, P.W. Form, function, and evolution in skulls and teeth of bats. In Bat Biology and Conservation; Kunz, T.H., Racey, P.A., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1998; pp. 140–156. ISBN 9781560988250. [Google Scholar]
- Schluter, D.; Nagel, L.M. Parallel speciation by natural selection. Am. Nat. 1995, 146, 292–301. [Google Scholar] [CrossRef]
- Muschick, M.; Indermaur, A.; Salzburger, W. Convergent evolution within an adaptive radiation of cichlid fishes. Curr. Biol. 2012, 22, 2362–2368. [Google Scholar] [CrossRef] [PubMed]
- Bonaccorso, F.J.; Winkelmann, J.R.; Shin, D.; Agrawal, C.I.; Aslami, N.; Bonney, C.; Hsu, A.; Jekielek, P.E.; Knox, A.K.; Kopach, S.J.; et al. Evidence for exploitative competition: Comparative foraging behavior and roosting ecology of short-tailed fruit bats (Phyllostomidae). Biotropica 2007, 39, 249–256. [Google Scholar] [CrossRef]
- Burbrink, F.T.; Pyron, R.A. How does ecological opportunity influence rates of speciation, extinction, and morphological diversification in New World ratsnakes (Tribe Lampropeltini)? Evolution 2009, 64, 934–943. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Shifts in adaptive peaks for jaw morphology of Neotropical frugivorous bats. Shifts in adaptive peaks are indicated with asterisks and convergent shifts in adaptive peaks have the same branch color.
Figure 1.
Shifts in adaptive peaks for jaw morphology of Neotropical frugivorous bats. Shifts in adaptive peaks are indicated with asterisks and convergent shifts in adaptive peaks have the same branch color.

Figure 2.
Shifts in adaptive peaks for skull morphology of Neotropical frugivorous bats. Shifts in adaptive peaks are indicated with asterisks and convergent shifts in adaptive peaks have the same branch color.
Figure 2.
Shifts in adaptive peaks for skull morphology of Neotropical frugivorous bats. Shifts in adaptive peaks are indicated with asterisks and convergent shifts in adaptive peaks have the same branch color.


{kind=link}
{kind=link}
Table 1.
Factor loads for the Phylogenetic Principal Component Analysis (pPCA) of morphometric variables of jaw and skull of frugivorous bats of Stenodermatinae and Carollinae subfamilies of Phyllostomidae. Total skull length (TSL), Postorbital Width (PW), Zygomatic Breadth (ZB), Posterior Skull width (PSW), Palatal Width at canines (PW), Palatal Width at first molar (PW1), Total Palate Length (TPL), Anterior Skull Length (ASL), Post-Palatal length (PPL), Maxillary Toothrow Length (MTL), Dentary Depth under first molar (DD), Coronoid Process Height (CPH), Condyle height (CH), Condyle to Canine bite point (CC), Condyle to first molar point (Cm1), Condyle to last molar point (Cm3), Total Dentary Length (TDL), Condyle-Coronoid Length (CCL), Coronoid-Angular Length (CAL) and Mandibular toothrow length (MAN).
Table 1.
Factor loads for the Phylogenetic Principal Component Analysis (pPCA) of morphometric variables of jaw and skull of frugivorous bats of Stenodermatinae and Carollinae subfamilies of Phyllostomidae. Total skull length (TSL), Postorbital Width (PW), Zygomatic Breadth (ZB), Posterior Skull width (PSW), Palatal Width at canines (PW), Palatal Width at first molar (PW1), Total Palate Length (TPL), Anterior Skull Length (ASL), Post-Palatal length (PPL), Maxillary Toothrow Length (MTL), Dentary Depth under first molar (DD), Coronoid Process Height (CPH), Condyle height (CH), Condyle to Canine bite point (CC), Condyle to first molar point (Cm1), Condyle to last molar point (Cm3), Total Dentary Length (TDL), Condyle-Coronoid Length (CCL), Coronoid-Angular Length (CAL) and Mandibular toothrow length (MAN).
| Trait | pPC1 | pPC2 | pPC3 | pPC4 | pPC5 |
|---|---|---|---|---|---|
| Jaw | |||||
| MDD | −0.850 | 0.519 | −0.041 | 0.049 | −0.066 |
| CPH | −0.990 | 0.112 | 0.045 | −0.013 | 0.071 |
| CH | −0.988 | −0.093 | 0.102 | 0.050 | −0.031 |
| CC | −0.997 | −0.041 | −0.064 | −0.013 | −0.006 |
| Cm1 | −0.996 | −0.043 | −0.058 | −0.035 | −0.006 |
| Cm3 | −0.988 | 0.021 | 0.039 | −0.142 | −0.034 |
| TDL | −0.995 | −0.065 | −0.073 | 0.004 | −0.029 |
| CCL | −0.998 | −0.014 | 0.019 | −0.012 | −0.001 |
| CAL | −0.995 | 0.016 | 0.084 | 0.014 | 0.019 |
| MAN | −0.989 | −0.043 | −0.129 | 0.042 | 0.024 |
| Skull | |||||
| TSL | −0.980 | −0.171 | −0.094 | 0.002 | −0.002 |
| MSW | −0.904 | −0.353 | −0.189 | −0.009 | −0.041 |
| ZB | −0.667 | −0.718 | −0.131 | −0.077 | −0.106 |
| PSW | −0.784 | −0.564 | −0.249 | −0.016 | −0.015 |
| PW | 0.118 | −0.988 | 0.077 | 0.069 | −0.010 |
| PW1 | −0.006 | −0.981 | 0.097 | −0.153 | 0.068 |
| TPL | −0.986 | 0.148 | 0.074 | 0.024 | 0.010 |
| ASL | −0.994 | −0.084 | −0.048 | 0.023 | 0.046 |
| PPL | −0.192 | 0.772 | −0.577 | 0.106 | 0.134 |
| MTL | −0.995 | −0.070 | 0.017 | −0.039 | −0.020 |
Table 2.
Wheatsheaf indices (w) with associated 95% confidence intervals for shape variables of jaw in each convergent regime. Wheatsheaf indices (w) with associated 95% confidence intervals for shape variables of skull in each convergent regime. p-values from analyses are also provided, and significant (p < 0.05) values are highlighted in bold. Factor loads indicated that pPC1 represented jaw size, pPC2 represented relationship between dentary depth, pPC3 represented relationship between condyle height and mandibular toothrow length, pPC4 represented condyle to last molar distance and pPC5 represented relations between dentary depth under first molar and coronoid process height.
Table 2.
Wheatsheaf indices (w) with associated 95% confidence intervals for shape variables of jaw in each convergent regime. Wheatsheaf indices (w) with associated 95% confidence intervals for shape variables of skull in each convergent regime. p-values from analyses are also provided, and significant (p < 0.05) values are highlighted in bold. Factor loads indicated that pPC1 represented jaw size, pPC2 represented relationship between dentary depth, pPC3 represented relationship between condyle height and mandibular toothrow length, pPC4 represented condyle to last molar distance and pPC5 represented relations between dentary depth under first molar and coronoid process height.
| Regimes | Phenotype | w | Lower Bound | Upper Bound | p-Value |
|---|---|---|---|---|---|
| 1 | pPC1 | 3.40 | 3.27 | 4.19 | 0.02 |
| pPC2 | 2.69 | 2.52 | 3.84 | 0.08 | |
| pPC3 | 4.08 | 3.96 | 5.44 | 0.04 | |
| pPC4 | 3.98 | 3.44 | 6.98 | 0.02 | |
| pPC5 | 3.65 | 3.51 | 5.04 | 0.06 | |
| 2 | pPC1 | 2.92 | 2.77 | 3.24 | <0.01 |
| pPC2 | 2.12 | 1.98 | 2.72 | 0.03 | |
| pPC3 | 4.84 | 4.59 | 6.10 | <0.01 | |
| pPC4 | 1.69 | 1.56 | 2.14 | 0.11 | |
| pPC5 | 1.73 | 1.62 | 1.92 | 0.14 | |
| 3 | pPC1 | 8.58 | 8.29 | 8.72 | 0.14 |
| pPC2 | 0.68 | 0.65 | 0.69 | 0.96 | |
| pPC3 | 3.37 | 3.27 | 3.41 | 0.36 | |
| pPC4 | 5.50 | 5.33 | 5.60 | 0.19 | |
| pPC5 | 10.28 | 9.87 | 10.45 | 0.20 | |
| 4 | pPC1 | 1.02 | 0.97 | 1.29 | 0.14 |
| pPC2 | 1.62 | 1.55 | 1.93 | 0.08 | |
| pPC3 | 1.81 | 1.72 | 2.01 | 0.01 | |
| pPC4 | 2.91 | 2.77 | 3.42 | <0.01 | |
| pPC5 | 1.31 | 1.26 | 1.57 | 0.17 |
Table 3.
Test and quantification of convergence, based on the Wheatsheaf index, of skull shape for convergent regimes of frugivorous bats of Stenodermatinae and Carollinae subfamilies of Phyllostomidae. Wheatsheaf indices (w) with associated 95% confidence intervals for shape variables of skull in each convergent regime. p-values from analyses are also provided, and significant (p < 0.05) values are highlighted in bold. pPC1 represented skull size, pPC2 represented relationship between palatal width and post-palatal length (PPL = 0.772), pPC3 represented post-palatal length, pPC4 represented relationship between palatal width and post-palatal length, and pPC5 represented relationship between zygomatic breadth and post-palatal length.
Table 3.
Test and quantification of convergence, based on the Wheatsheaf index, of skull shape for convergent regimes of frugivorous bats of Stenodermatinae and Carollinae subfamilies of Phyllostomidae. Wheatsheaf indices (w) with associated 95% confidence intervals for shape variables of skull in each convergent regime. p-values from analyses are also provided, and significant (p < 0.05) values are highlighted in bold. pPC1 represented skull size, pPC2 represented relationship between palatal width and post-palatal length (PPL = 0.772), pPC3 represented post-palatal length, pPC4 represented relationship between palatal width and post-palatal length, and pPC5 represented relationship between zygomatic breadth and post-palatal length.
| Regimes | Phenotype | w | Lower Bound | Upper Bound | p-Value |
|---|---|---|---|---|---|
| 1 | pPC1 | 2.57 | 2.42 | 3.98 | 0.06 |
| pPC2 | 4.95 | 4.71 | 5.86 | 0.03 | |
| pPC3 | 2.04 | 1.96 | 2.56 | 0.12 | |
| pPC4 | 2.37 | 2.27 | 4.10 | 0.12 | |
| pPC5 | 4.14 | 4.01 | 7.27 | 0.03 | |
| 2 | pPC1 | 1.90 | 1.79 | 2.57 | 0.03 |
| pPC2 | 5.08 | 4.76 | 5.69 | 0.01 | |
| pPC3 | 2.24 | 2.11 | 2.28 | 0.03 | |
| pPC4 | 1.57 | 1.45 | 2.01 | 0.05 | |
| pPC5 | 1.00 | 0.96 | 1.06 | 0.44 | |
| 3 | pPC1 | 0.99 | 0.93 | 1.15 | 0.18 |
| pPC2 | 0.91 | 0.87 | 1.04 | 0.35 | |
| pPC3 | 2.94 | 2.79 | 3.56 | <0.01 | |
| pPC4 | 2.29 | 2.03 | 2.56 | 0.01 | |
| pPC5 | 1.30 | 1.24 | 1.49 | 0.05 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Murillo-García, O.E.; De la vega, M.E. Divergence, Convergence and Phenotypic Diversity of Neotropical Frugivorous Bats. Diversity 2018, 10, 100. https://doi.org/10.3390/d10030100
AMA Style
Murillo-García OE, De la vega ME. Divergence, Convergence and Phenotypic Diversity of Neotropical Frugivorous Bats. Diversity. 2018; 10(3):100. https://doi.org/10.3390/d10030100
Chicago/Turabian StyleMurillo-García, Oscar E., and Maria E. De la vega. 2018. "Divergence, Convergence and Phenotypic Diversity of Neotropical Frugivorous Bats" Diversity 10, no. 3: 100. https://doi.org/10.3390/d10030100
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.