Kuschelysius, a New Alpine Genus of Eugnomine Weevil (Coleoptera: Curculionidae: Curculioninae) from New Zealand


1
The New Zealand Institute for Plant and Food Research Ltd., Private Bag 92169, Auckland Mail Centre, Auckland 1142, New Zealand
2
New Zealand Arthropod Collection, Manaaki Whenua Landcare Research, Private Bag 92170, Auckland 1142, New Zealand
*
Author to whom correspondence should be addressed.
Diversity 2018, 10(3), 75; https://doi.org/10.3390/d10030075
Submission received: 27 June 2018
/
Revised: 22 July 2018
/
Accepted: 24 July 2018
/
Published: 27 July 2018
(This article belongs to the Special Issue Systematics and Phylogeny of Weevils)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Kuschelysius new genus is described for four species, K. hollowayae new species, K. durus new species, K. verbalis new species and K. nitens new species, which are found in alpine regions along the length of the South Island of New Zealand. The genus most closely resembles members of the genus Eugnomus but is distinguished from them by the presence of a small pair of post-ocular tubercles and by having appressed scales on the dorsal surfaces. Some members of Kuschelysius appear to be flight-capable with well-developed hindwings, while others have reduced hindwings and are presumably flightless. Many specimens have been collected from the flowers of Dracophyllum traversii, Celmisia and other alpine plants, and the guts of examined specimens contained pollen. We hypothesise that the species of Kuschelysius are pollinators of the New Zealand alpine flora.
1. Introduction
The Eugnomini Lacordaire, 1863 are a tribe of curculionine weevils with a primarily Southern Hemisphere distribution. The tribe is present in Australia, New Zealand, New Caledonia, South America extending northward to Central America, with undescribed members also known from Indochina [1]. The Eugnomini reach their greatest diversity in New Zealand, with approximately 100 species in 19 endemic genera, one genus (Rhopalomerus Blanchard) shared with Australia and South America and another (Pactola Pascoe) shared with New Caledonia and Fiji [2]. They form a distinctive element of the New Zealand weevil fauna, where their diurnal activity, often bizarre morphologies masquerading as faeces while feigning death, and flower-frequenting behaviour [3,4] attract the attention of natural historians.
The exact composition of the Eugnomini remains unclear. A suite of characters has been used to characterise the tribe. These characters include an elongate head with the eyes separated from the head constriction by a distance greater than their own length, scrobes that run below the lower surface of the rostrum, front coxae conical, and a large, obvious tooth on the hind tibia [3]: some of these characters are not universally shared and there have not yet been any phylogenetic studies to identify synapomorphies that unequivocally indicate the monophyletic nature of the tribe [1]. Substantial advances have been made in recent years on the eugnomine weevil fauna of New Caledonia and the Pacific [2,5,6,7,8,9], but the remainder of the Eugnomini of the world remain understudied.
Despite the lack of regional or global revisions of the tribe, the lowland eugnomine weevils of New Zealand are reasonably well known and have been named and inventoried through Broun’s pioneering taxonomic research [10,11,12,13,14], illustrated accounts of several species by Hudson [15,16] and a key to genera by Marshall [17]. However, it is becoming apparent that the diversity of weevils in alpine areas has yet to be fully appreciated, with many species belonging to several genera being found exclusively above the treeline [18].
This contribution describes a new genus of eugnomine weevil containing four distinctive, large, alpine species, which we dedicate to our late friend, mentor and colleague Guillermo (Willy) Kuschel. We also name the type species after our friend and colleague Beverly Holloway, Willy’s wife, a leading coleopterist in her own right. Although Willy’s published contributions on the Eugnomini were only ever part of broader works [19,20,21,22], he was fascinated by the group and worked on the fauna of New Zealand during his first lengthy visit to the country and soon after his permanent settlement in New Zealand. Regrettably, the results from this research were not published in his lifetime.
2. Materials and Methods
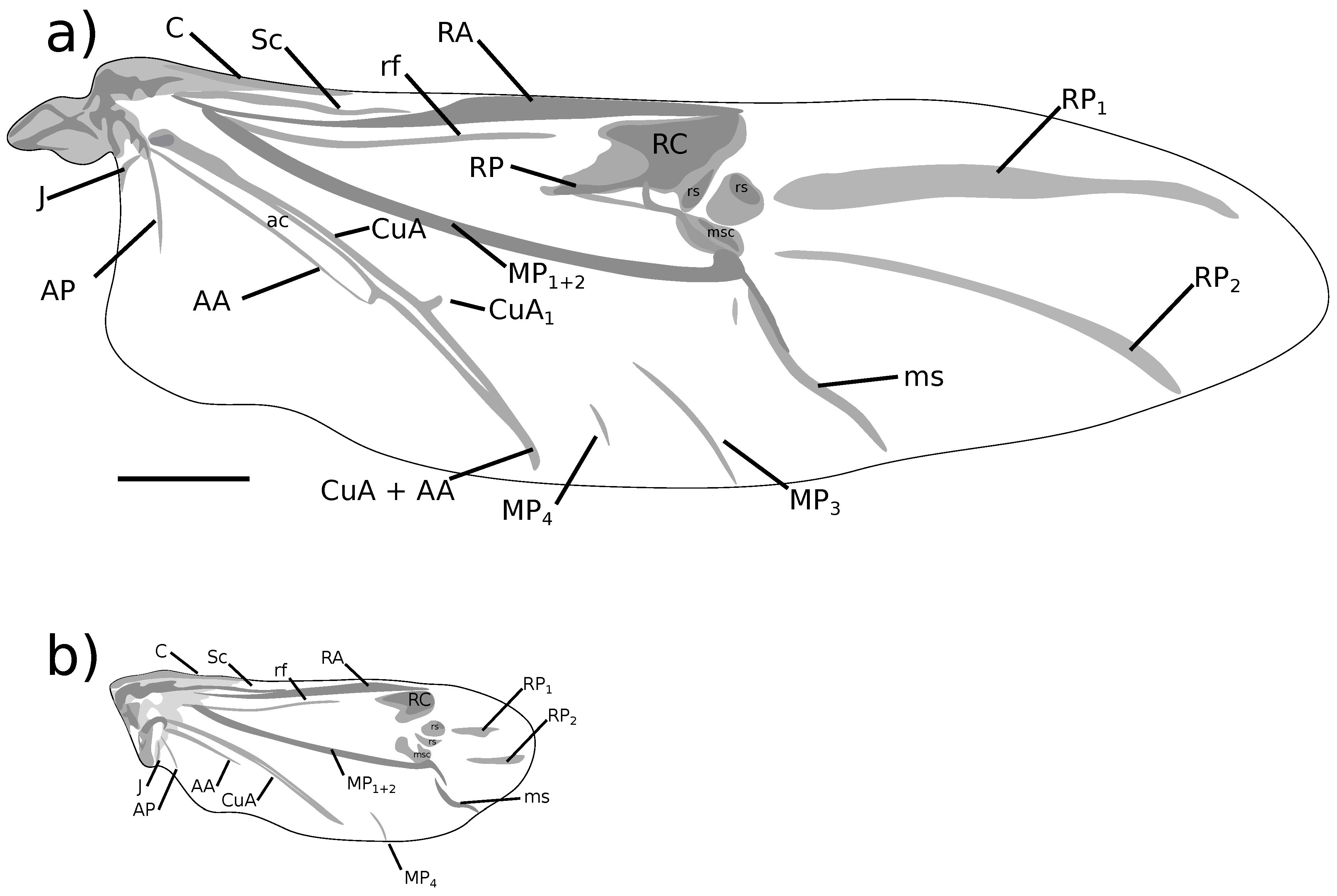
Morphological features are described using standard terminology [23,24,25], with the following additions. Body length was measured in lateral view, from the anterior margin of the eyes to the elytral declivity. Body height was measured in lateral view as a straight line from the hind coxae to the dorsal surface of the elytra. Rostrum width was measured across the antennal insertions. The curve formed by the connection of the head and rostrum, as observed in lateral view, is termed the “ventral curvature of the head”. The degree of curvature can be described as “gently curved” (i.e., having a large circle of curvature) or “tightly curved” (i.e., having a small circle of curvature). Wing venation terminology follows Kukalová-Peck and Lawrence [26], as applied to Curculionoidea by Oberprieler et al. [23]. This relates to the terminology of Zherikhin and Gratshev [27] in the following way: C = C; Sc = Sc; RA = R; RC = w; rf = rf; RP = Rr; RP = pst; RP = h & mst; rs = rs; MP = Cu; msc = msc; ms = af; MP = 1A; MP = 1A; CuA = 2A; CuA = a–a AA = 3A; ac = ac; AP = 4A; J = J.
Descriptions of colour follow the terminology provided by the National Bureau of Standards [28], which gives 267 centroid colours with natural-language descriptions. Digital representations of these colours have been provided by Jaffer [29].
Genitalia were photographed in KY Jelly (Johnson & Johnson Pacific, Broadway, NSW, Australia) before being stored in glycerol in a vial pinned below the specimen. Illustrations were prepared from photographs, using Inkscape (v. 0.91, [30]). Habitus photographs were taken using Nikon DS-Ri1 (Melville, NY, USA) fitted with a digital camera and a mechanical z-stepper. Nikon NIS Elements v. 4.10 was used to prepare the image stack and to produced the final montaged image.
Specimens were examined and deposited in the following collections:
- FRNZ:
- The National Forest Insect Collection, Scion, Rotorua, New Zealand
- LUNZ:
- Lincoln University Entomology Research Museum, Lincoln, Canterbury, New Zealand
- NHM:
- Natural History Museum, London, United Kingdom
- NZAC:
- New Zealand Arthropod Collection, Manaaki Whenua Landcare Research, Tamaki, Auckland, New Zealand
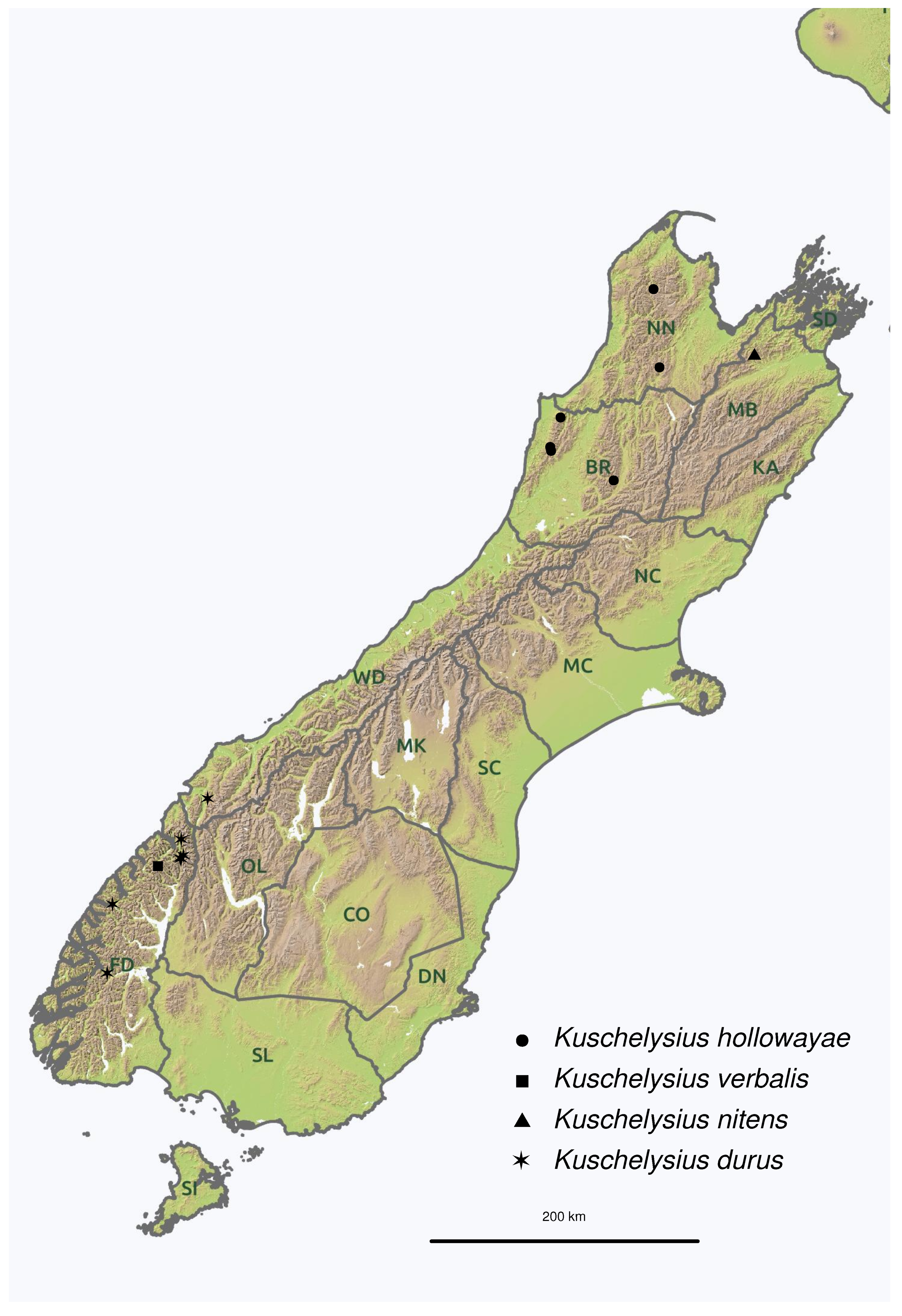
Label data from holotypes are transcribed using the following conventions. Data from individual labels are enclosed using quotes (‘…’), lines are indicated with a solidus (/) and metadata are given in square brackets ([…]). Two-letter regional codes (NN, BR, FD, etc., Figure 1) follow those proposed by Crosby et al. [31].
3. Results
3.1. Kuschelysius Brown and Leschen New Genus
Type species: Kuschelysius hollowayae Brown and Leschen new species, by present designation.
3.1.1. Diagnosis
Large-bodied (body length > 5.0 mm, height > 1.7 mm, width > 2.2 mm); dorsal surfaces covered with appressed scales; antennal club with segment 3 shorter than segments 1 and 2 combined; length of the rostrum longer than the head; head not constricted behind eyes, tubercles present behind eyes; distinct spines absent from pronotum and elytra; elytra with a humeral callus; all femora with a single ventral tooth; all tibiae sinuous; tarsal claws simple, lacking a tooth.
The presence of a pair of small tubercles present behind the eyes will distinguish Kuschelysius from most other genera of New Zealand eugnomine weevils. Kuschelysius is most similar to species of Eugnomus Schönherr: the presence of appressed scales which conceal the integument, and sinuous pro- and mesotibiae distinguish them from Eugnomus, which have dorsal vestiture of fine hairs that reveal the integument, head evenly convex behind the eyes and straight pro- and mesotibiae. Tysius Pascoe also has tubercles behind the eyes; however, this genus can be distinguished from Kuschelysius by its much smaller size (body length < 3 mm), eyes placed further onto the rostrum, round scutellar shield and by the elytra having facia on interstria 3 and a low tubercle on interstria 5 on the elytral declivity.
3.1.2. Description
Rostrum. Mandibles stout, not exodont. Maxillae with long and flexible palps. Antennae inserted laterally, at distal 1/4 of rostrum. Scrobes oblique, running along ventral surface for 2/3 length, terminating just short of eyes. Head. Eyes hemispherical, prominent, positioned anterior of point of maximum ventral head curvature. Head not constricted behind eyes. Tubercles present behind eyes. In lateral view, angle formed by ventral margin of rostrum and head capsule c. 130°. Antennae. Scape reaching posterior margin of eyes when in repose, resting position running along ventral margin of rostrum. Funicle with 7 segments. Segments stout, clothed with dense thick setae; segments 1 and 2 lengths subequal, each about as long as 3 and 4 combined. Pronotum. Widest posteriorly, about 3/5 as wide as combined width of elytra; width at anterior margin much narrower than width at posterior margin. Lateral margins constricted in anterior 1/4 before abruptly widening, subparallel in posterior 2/3. Scales on disc larger than those on the elytra. Elytra. Stria 10 complete. Humeral callus developed. Disc without tubercles or spines. Wings. Reduced to fully developed. Costal margin straight, apex widely rounded. RA strongly sclerotised, and widest around middle. Radial cell (RC) completely sclerotised. RP clearly evident. MP wide and strongly sclerotised. CuA wide at base, divided in middle to form a long and narrow pseudocell. AP short, not reaching wing margin. Thoracic ventrites. Prosternum projecting ventrally, resulting in it having an anterior face. Mesoventral process swollen. Legs. All femora armed with a single ventral tooth; metafemoral tooth large, not excised at base of distal edge. All tibiae sinuous. Tarsal segment 1 stout, shorter than combined length of remaining segments; segment 5 about 1.5 times as long as segment 3. Male genitalia. Pedon tubular, relatively short, broad and high; membranous ventrally; base of pedon with a narrow, strongly sclerotised ventral brace. Temones approximately as long as pedon. Parameroid lobes elongate, fused along proximal 1/2. Manubrium stout, shorter than temones. Spiculum gastrale with furcal arms very broad, maximum width approximately 0.5 times length of apodeme. Female genitalia. Styli slender, inserted on ventral margin of gonocoxites. Gonocoxites short and broad. Bursa copulatrix long, apparently with two chambers. Sternite 8 entire, apex broadly rounded. Spermatheca C-shaped, slender.
3.1.3. Etymology
Named after Dr Guillermo Kuschel Gerdes (1918–2017) whose research into the weevils of the Southern Hemisphere gave substantial insight into weevil classification. The ending ‘elysius’ refers to Elysium, the ‘Land of Joy’ of Antiquity, which was located by Plato at the antipodes [32].
3.2. Kuschelysius hollowayae Brown and Leschen New Species
3.2.1. Diagnosis
Uniformly light greyish brown, vestiture largely decumbent. Elytra long, 1.75 times longer than wide and 3.6 times longer than pronotum; declivity gently sloping; elytral apices square. Profemoral tooth small.
3.2.2. Description
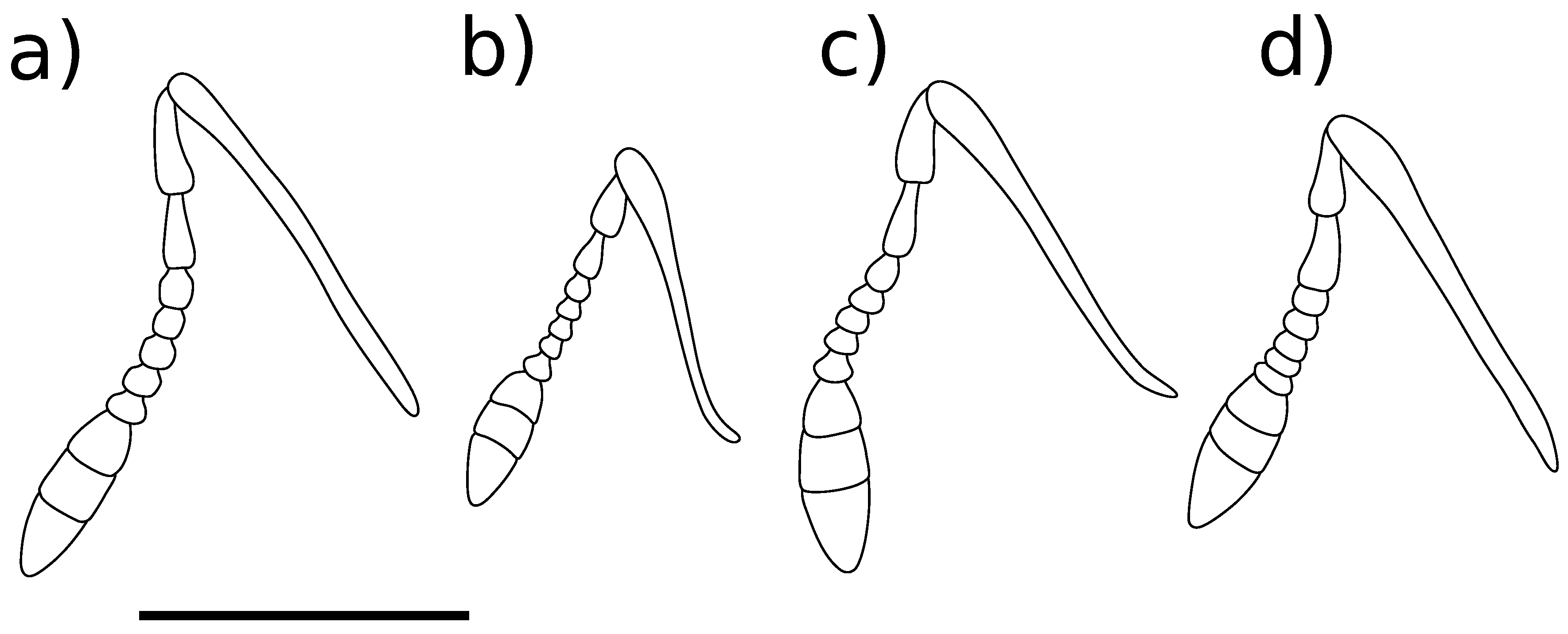
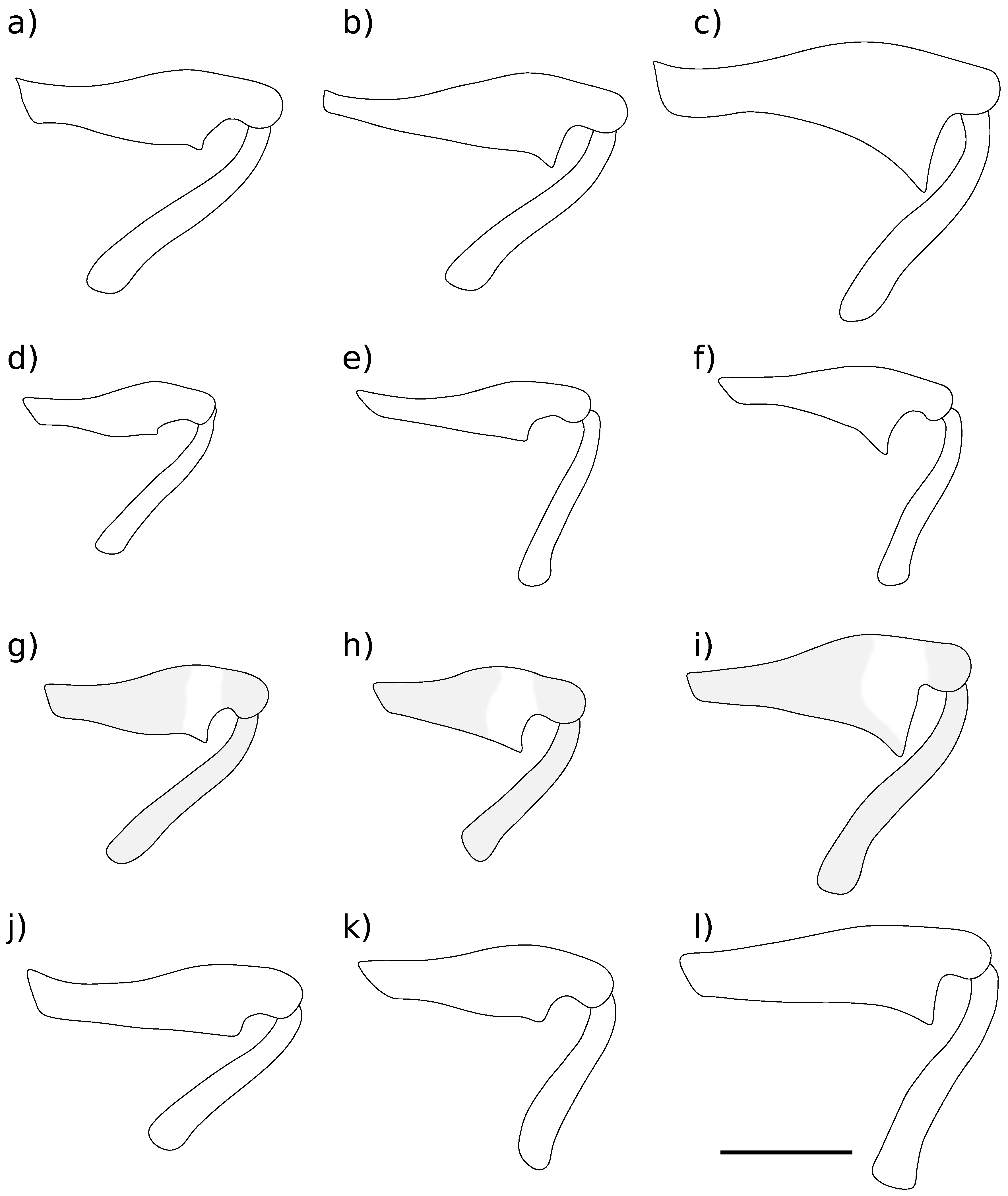
Body length 7.22 mm to 8.05 mm ( = 7.80 mm, s = 0.39, n = 4), height 2.46 mm to 2.65 mm ( = 2.57 mm, s = 0.08, n = 4). Integument reddish black. Pronotum and elytra covered with adpressed light greyish brown scales and sparse, erect, trichiform setae. Scales present on head, rostrum, procoxae, mesoventrite and distal 1/3 of tibiae, paler than scales on dorsum. Rostrum. Length 1.78 mm to 1.98 mm ( = 1.88 mm, s = 0.09, n = 4), width 0.49 mm to 0.59 mm ( = 0.54 mm, s = 0.04, n = 4), length/width ratio 3.12 to 3.67 ( = 3.48, s = 0.25, n = 4). Straight in lateral view. Dorsal surface with elongate, matte scales. Antennae. Figure 4a. Scape clothed with scales and setae. Funicle segments 3–6 subequal, approximately as long as wide, segment 7 transverse. Head. Postocular tubercles evident. Ventral curvature of head tightly curved. Pronotum. Length 1.56 mm to 1.70 mm ( = 1.63 mm, s = 0.06, n = 4), width 1.60 mm to 1.77 mm ( = 1.71 mm, s = 0.07, n = 4), length/width ratio 0.93 to 0.98 ( = 0.96, s = 0.02, n = 4). Lateral margins moderately constricted in dorsal view in anterior 1/4. Scales on disc matte, sparser than elytral scales. Scutellar shield square. Elytra. Length 5.61 mm to 5.99 mm ( = 5.81 mm, s = 0.18, n = 4), width 3.04 mm to 3.54 mm ( = 3.33 mm, s = 0.23, n = 4), length/width ratio 1.63 to 1.85 ( = 1.75, s = 0.10, n = 4). Lateral margins subparallel in anterior 2/3 in dorsal view, tapering toward apex in posterior 1/3. Scales smaller, more elongate, denser than on pronotum. All interstriae evenly convex. Elytral declivity gently curved in lateral view. Apices of each elytron square. Wings. Figure 3a. Fully developed, 9.34 mm long from apex to jugal margin, 3.01 mm wide at midpoint (n = 1). MP long, touching wing margin; MP much shorter, c. 1/3 length of MP, not touching wing margin. AA parallel to CuA along most of length, becomes weak and inconspicuous before joining CuA proximally to the branching of CuA. Thoracic ventrites. Mesoventrite densely clothed with pale scales, contrasting with sparser vestiture on mesanepisternum. Mesoventral projection narrowly rounded at apex. Abdomen. Tergite 7 wider than long in males; longer than wide, with rounded apex in females. Tergite 8 subquadrate, exposed in males (Figure 6a); strongly arched, bifurcate with acute apices, concealed under tergite 7 in females (Figure 6f,g). Ventrite 5 depressed medioapically in males; raised medioapically in females. Legs. Profemoral tooth small (Figure 5a). Mesofemoral tooth moderate (Figure 5b). Metafemoral tooth large (Figure 5c). Male genitalia. Figure 6a–e. Pedon with lateral lobes separated dorsally. Internal sac armed with small teeth, localised into elongate regions dorsally and ventrally. Female genitalia. Figure 6h–j. Apices of styli with a long flexible seta. Gonocoxites in lateral view high in the basal 7/8, rapidly narrowing to styli.
3.2.3. Holotype
Male (NZAC). Specimen mounted on card triangle; left metanepisternite partially dissociated; abdomen removed for dissection, stored in glass vial pinned below specimen. Labelled: ‘Mt. Owen 5000’/Nelson, 21.10.62/J.I. Townsend’ [first two lines handwritten, last line printed, rectangular off white paper], ‘Celmisia/armstrongii’ [printed, rectangular off white paper], [narrow strip of green paper], ‘HOLOTYPE/Kuschelysius/hollowayae/Brown & Leschen 2018’ [printed, red card].
3.2.4. Paratypes
A total of 42 specimens (31 males, 10 females, 1 of undetermined sex) designated as paratypes, bearing blue paratype label. Paratype specimens deposited in NZAC, LUNZ, NHM, FRNZ.
NN. Mt. Owen 5000’, 21 Oct 1962, JI Townsend, Celmisia armstrongii (NZAC: 8); Mt. Owen 5000’, 30 Dec 1962, JI Townsend (NZAC: 2); Mt. Owen, 12 Jan 1962, JS Dugdale (NZAC: 2); Mt. Owen, 7 Nov 1961, D Kershaw (FRNZ: 1); Mt. Aorere, 5 Dec 1962, W.A.H. (FRNZ: 1). BR. Paparoa Range, Mt. Dewar 1100 m, Dec 1969, JI Townsend, Dracophyllum traversii flower heads (NZAC: 13, NHM: 4); Paparoa Range, Mt. Dewar 1060 m, Dec 1969, JG McBurney (NZAC: 1); Paparoa Range, Lochnagar Ridge 3500–3800’, 2–10 Dec 1969, JS Dugdale (NZAC: 1); Paparoa Range, Lochnagar Ridge 1067 m, Dec 1969, JS Dugdale and JI Townsend, Dracophyllum traversii (NZAC: 6); Paparoa Range, Lochnagar Ridge, camp area 1060 m, Dec 1969, JI Townsend (NZAC: 1); Paparoa Range, Buckland Peaks, Townson Tarn 1200 m, 15 Nov 1987, BP Stephenson, under rocks (LUNZ: 1); Victoria Range, south end, head of Rahu Creek trib., 16 Jan 1967, JS Dugdale (NZAC: 1).
3.2.5. Distribution
South Island, NN, BR: Domett Range, Mount Owen, Paparoa Range, Victoria Range (Figure 1).
3.2.6. Biology
This species has frequently been collected on the flowerheads of Dracophyllum traversii Hook. f., and less commonly on Celmisia armstrongii Petrie. Further research is required to ascertain whether these records are an accurate indication of the larval host plant, or if the adults were only incidentally feeding on these plants. Adults have been collected at elevations between 1060 and 1520 m above sea level. Larvae are currently unknown.
3.2.7. Etymology
Named after Dr Beverley Holloway, Willy Kuschel’s wife of over 50 years. She has made significant contributions to the taxonomy of the Anthribidae and the Lucanidae and is a fellow of the Entomological Society of New Zealand.
3.3. Kuschelysius durus Brown and Leschen New Species
3.3.1. Diagnosis
Evenly yellowish grey; small dark greyish brown maculae on elytra. Elytra long, 1.7 times longer than wide and 3.2 times longer than pronotum. Profemoral tooth small.
3.3.2. Description
Body length 5.39 mm to 5.86 mm ( = 5.64 mm, s = 0.21, n = 4), height 1.74 mm to 2.24 mm ( = 1.98 mm, s = 0.21, n = 4). Integument reddish to black. Pronotum and elytra covered with appressed yellowish grey scales and sparse, erect trichiform setae; small dark greyish brown maculae present on the elytra, position variable but often with two maculae present on interstria 3 on disc and at top of elytral declivity. Thoracic ventrites clothed with yellowish grey scales; abdominal ventrites with short hairs. Rostrum. Length 1.12 mm to 1.21 mm ( = 1.16 mm, s = 0.04, n = 4), width 0.40 mm to 0.42 mm ( = 0.41 mm, s = 0.01, n = 4), length/width ratio 2.73 to 2.88 ( = 2.82, s = 0.07, n = 4). Straight in lateral view. Dorsal surface with oval, matte scales. Antennae. Figure 4b. Scape clothed with setae only. Funicle segments 3–6 subequal, approximately as long as wide, segment 7 transverse. Head. Postocular tubercles reduced. Ventral curvature of head tightly curved. Pronotum. Length 1.14 mm to 1.38 mm ( = 1.27 mm, s = 0.10, n = 4), width 1.19 mm to 1.36 mm ( = 1.28 mm, s = 0.08, n = 4), length/width ratio 0.96 to 1.02 ( = 1.00, s = 0.03, n = 4). Lateral margins moderately constricted in dorsal view in anterior 1/4. Scales on disc overlapping, matte, larger but about as dense as scales on elytra. Scutellar shield pentagonal. Elytra. Length 3.85 mm to 4.26 mm ( = 4.04 mm, s = 0.20, n = 4), width 2.22 mm to 2.59 mm ( = 2.42 mm, s = 0.16, n = 4), length/width ratio 1.63 to 1.75 ( = 1.67, s = 0.06, n = 4). All interstriae evenly convex. Scales on disc overlapping, matte, about as dense as scales on pronotum. Elytral declivity gently curved in lateral view. Apices square. Wings. Figure 3b. Reduced, particularly with respect to the apical sector, 3.05 mm to 3.08 mm long from apex to jugal margin, 1.03 mm to 1.21 mm wide at midpoint (n = 2). RP and RP present, but much reduced in length. MP absent; MP touching wing margin. AA parallel to CuA, no evidence of it joining CuA. CuA absent. Thoracic ventrites. Mesoventrite, mesanepisternum, metanepisternum and lateral 1/4 of the first two abdominal ventrites densely clothed with pale yellow scales, contrasting with brilliant yellow hairs on disc of metaventrite. Mesoventral projection truncate at apex. Abdomen. Tergite 7 wider than long in males; elongate with rounded apex in females. Tergite 8 subquadrate, exposed in males (Figure 7a); weakly arched, bifurcate with blunt apices, concealed under tergite 7 in females (Figure 7f). Ventrite 5 depressed medioapically in males; raised medioapically in females. Legs. Profemoral tooth small (Figure 5d). Mesofemoral tooth small (Figure 5e). Metafemoral tooth large, acute (Figure 5f). Male genitalia. Figure 7a–e. Pedon with lateral lobes dorsally fused in basal 1/4, with a lobe projecting apicad. Internal sac armed with two large, toothed sclerites. Female genitalia. Figure 7f–j. Apices of styli with multiple long setae. Gonocoxites in lateral view of roughly even height along length, gradually narrowing to styli.
3.3.3. Holotype
Female (NZAC). Specimen glued onto card mount; entire. Labelled: ‘Gertrude Saddle’/Homer 5.2.63/R.M.Bull’ [handwritten, rectangular off white paper], ‘R.M. Bull/Collection’ [printed, rectangular white card], ‘HOLOTYPE/Kuschelysius/durus/Brown & Leschen 2018’ [printed, red card].
3.3.4. Paratypes
A total of 17 specimens (6 males, 11 females) designated as paratypes, bearing blue paratype label. Paratype specimens deposited in NZAC, NHM.
WD. Annetta Mountain, Barrier Valley, 2 Feb 1975, JS Dugdale, in turf (NZAC: 1). FD. Mt. Titiroa, Borland side W, 6 Feb 2009, R Hoare, On cushion plant gravel field 45.04° S 167.31° E (NZAC: 1); Gertrude Saddle, Homer, 4500’, 5 Feb 1963, JI Townsend, Astelia nivicola (NZAC: 6, NHM: 2); Gertrude Saddle, Homer, 4500’, 5 Feb 1963, JI Townsend, Celmisia (NZAC: 1); Homer Tunnel, 3000’, 5 Feb 1963, RM Bull (NZAC: 2); South Basin, Tutoko Bench, Darran Mt.s, 1219–1372 m, 15 Jan 1977, JS Dugdale (NZAC: 1); Homer Saddle, 4200’, 29 Jan 1946, R Forster, in leafmould (NZAC: 1); Mt. Barber, 1350 m, Jan 1970, J Dugdale, Celmisia walkeri (NZAC: 2).
3.3.5. Distribution
South Island, WD: Annetta Mountain. FD: Darran Mountains, Homer Tunnel, Mount Titiroa, Mount Barber (Figure 1).
3.3.6. Biology
This species has been collected in leaf litter, in turf, on unidentified cushion plants and on Celmisia walkeri Kirk. A large series of specimens have been collected from Astelia nivicola Ckn. ex Cheesm. The gut contents of the two dissected specimens collected from A. nivicola were filled almost exclusively with a single form of pollen. The specimen from Mt. Titiroa was collected in an unusual granite sand plain ecosystem [33,34]. Adults have been collected at elevations between 900 and 1380 m above sea level. Larvae are currently unknown.
3.3.7. Etymology
Based on the Latin durus, ‘strong, tough’, in reference to the hardiness of this species which survives in the harsh environment of the Fiordland mountains. It is also a trait demonstrated by Willy, whose endurance while undertaking collecting expeditions in the Juan Fernandez Islands, New Zealand and elsewhere in the South Pacific is awe-inspiring.
3.4. Kuschelysius verbalis Brown and Leschen New Species
3.4.1. Diagnosis
Mottled dark olive brown and greyish yellow. Mottling on elytra not forming any particular pattern, but forming paired dorsal vittae on the pronotum and broad bands on the distal 1/4 of the femora. Elytra relatively short, 1.5 times longer than wide and 3.2 times longer than pronotum. Profemoral tooth large (Figure 5g).
3.4.2. Description
Body length 6.20 mm (n = 1), height 2.38 mm (n = 1). Integument black; all surfaces covered with oval scales, mainly coloured dark olive brown but greyish yellow scales forming a ventral stripe below and posterior maculae above the eyes, paired dorsal vittae on the pronotum, a mottled pattern on the elytra, a patch on the anterior surface of the prosternum and broad bands on the distal 1/4 of the femora. Sparse, erect trichiform setae present on the rostrum, pronotum and elytra. Rostrum. Length 1.76 mm (n = 1), width 0.48 mm (n = 1), length/width ratio 3.67 (n = 1). Evenly curved in lateral view. Dorsal surface with oval, matte scales. Antennae. Figure 4c. Scape clothed with setae only. Funicle segment 3 longer than wide; segments 4–6 subequal, approximately as long as wide, segment 7 transverse. Head. Postocular tubercles reduced. Ventral curvature of head gently curved. Pronotum. Length 1.37 mm (n = 1), width 1.41 mm (n = 1), length/width ratio 0.97 (n = 1). Lateral margins strongly constricted in dorsal view in anterior 1/4. Scales on disc overlapping, matte, about as dense as scales on elytra. Scutellar shield square. Elytra. Length 4.37 mm (n = 1), width 2.92 mm (n = 1), length/width ratio 1.50 (n = 1). Interstriae 3 and 5 raised around the elytral declivity. Scales on disc overlapping, matte, about as dense as scales on pronotum. Elytral declivity evenly sloped in lateral view. Apices square. Thoracic ventrites. Densely clothed with scales of similar colour to dorsum, but longer, thinner and with different texture. Mesoventral projection truncate at apex. Abdomen. Ventrite 5 of females swollen medially. Legs. Profemoral tooth moderate (Figure 5g). Mesofemoral tooth moderate (Figure 5g). Metafemoral tooth large (Figure 5i). Male genitalia. Unknown. Female genitalia. Not examined.
3.4.3. Holotype
Female (NZAC). Specimen mounted on card triangle; entire. Labelled: ‘MacKinnan[sic] Pass 3500’/1.1.63 B.M.May/On Celmisia sp.’ (handwritten, rectangular off white paper, ‘Celmisia’ underlined), ‘HOLOTYPE/Kuschelysius/verbalis/Brown & Leschen 2018’ [printed, red card].
3.4.4. Distribution
South Island, FD: Mackinnon Pass (Figure 1).
3.4.5. Etymology
Based on the Latin verbalis, ‘of words,’ an allusion to Willy’s enjoyment of language.
3.5. Kuschelysius nitens Brown and Leschen New Species
3.5.1. Diagnosis
Uniformly medium grey; pronotum, elytra and legs with fine pale yellow-green setae that project over ground vestiture. Elytra long, 1.68 times longer than wide and 3.9 times longer than pronotum; declivity rounded in lateral view; elytral apices individually rounded. Profemoral tooth small.
3.5.2. Description
Body length 7.79 mm (n = 1), height 2.96 mm (n = 1). Integument black; rostrum, pronotum, elytra, legs and venter covered with fine, glossy adpressed medium grey scales and sparse, decumbent, pale yellow-green trichiform setae. Rostrum. Length 1.82 mm (n = 1), width 0.55 mm (n = 1), length/width ratio 3.31 (n = 1). Straight in lateral view. Dorsal surface with oval, glossy scales. Antennae. Figure 4d. Scape clothed with scales and setae. Funicle segments 3–5 subequal, approximately as long as wide, segments 6 and 7 transverse. Head. Postocular tubercles reduced. Ventral curvature of head tightly curved. Pronotum. Length 1.51 mm (n = 1), width 1.81 mm (n = 1), length/width ratio 0.83 (n = 1). Lateral margins moderately constricted in dorsal view in anterior 1/4. Scales on disc oval, glossy, about as dense as scales on elytra. Scutellar shield square. Elytra. Length 5.95 mm (n = 1), width 3.55 mm (n = 1), length/width ratio 1.68 (n = 1). All interstriae evenly convex. Scales on disc oval, glossy, about as dense as scales on pronotum but smaller. Elytral declivity strongly curved in lateral view. Apices of each elytron individually rounded. Thoracic ventrites. Mesoventral projection rounded at apex. Legs. Profemoral tooth small, broadly rounded (Figure 5j). Mesofemoral tooth small (Figure 5k). Metafemoral tooth moderate (Figure 5l). Male genitalia. Not examined. Female genitalia. Unknown.
3.5.3. Holotype
Male (NZAC). Specimen mounted on card triangle; entire, head bent to the left. Labelled: ‘Mt. Richmond/5000 23.4.63/G. Ramsay’ [handwritten, rectangular off white paper], ‘HOLOTYPE/Kuschelysius/nitens/Brown & Leschen 2018’ [printed, red card].
3.5.4. Distribution
South Island, MB: Mt. Richmond (Figure 1).
3.5.5. Etymology
Based on the Latin nitens ‘shining,’ in reference to the glossy scales that distinguish this species from others in the genus.
3.5.6. Remarks
Four New Zealand peaks are named Mt. Richmond. The one identified as the type locality is Mt. Richmond in the Richmond Range (MB; 41.4744° S 173.3957° E). This peak is readily accessible from Nelson, where G.W. Ramsay was living at the time of the collection of this specimen. Its maximum elevation is 1760 m (=5800 feet), which is consistent with the elevation inferred from the label (5000 feet = 1524 m).
The alternatives include Mt. Richmond, in the Southern Alps north of Lake Tekapo (MK; 43.5294° S 170.4314° E, 2509 m), which is flanked by glaciers and with very steep gradients around the elevation of interest; Mt. Richmond in the Livingstone Mountains, west of North Mavora Lake (OL; 45.2331° S 168.1110° E, 1673 m) which is of appropriate elevation but is poorly accessible; or Ōtāhuhu/Mount Richmond on the Auckland isthmus (AK; 36.9339° S 174.8385° E, 48 m) which is too low to be the type locality.
4. Key to Species of Kuschelysius
- 1.
- Dorsum mottled darker and lighter brown, without any clear pattern. Rostrum greater than 3.5 times longer than wide. Elytra around 1.5 times longer than wide. Profemoral tooth large, acute . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . K. verbalis
- -
- Dorsum uniformly coloured, brown or grey, or yellowish grey with small dark brown spots on interstriae 3 and 5. Rostrum less than 3.5 times longer than wide. Elytra greater than 1.6 times longer than wide. Profemoral tooth small, broadly rounded . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2.
- 2
- (1). Body length less than 6 mm. Scape clothed with setae only . . . . . . . . . . . . . . . . . . . . . K. durus
- -
- Body length greater than 7 mm. Scape clothed with appressed scales as well as setae . . . . . . . . 3.
- 3
- (2). Scales on pronotum and elytra matte. Elytral declivity gently sloping in lateral view, apex pointed. Setae on pronotum and elytra erect, dark . . . . . . . . . . . . . . . . . . . . . . . . . . . . K. hollowayae
- -
- Scales on pronotum and elytra glossy. Elytral declivity rounded in lateral view, apex rounded. Setae on pronotum and elytra decumbent, pale yellow-green . . . . . . . . . . . . . . . . . . . . . . K. nitens
5. Discussion
Although there have been several surveys of alpine insects in New Zealand [34,35], investigations into the evolutionary history of these taxa and the high level of endemism in these environments are still in their infancy [36]. The alpine beetle fauna is quite rich, and almost every mountain range in the Southern Alps is home to endemic species [18,37]. The New Zealand Eugnominae are common in alpine environments, with four other genera (Eugnomus Schönherr, 1847, Oreocalus May 1993, Pactolotypus Broun, 1909, Stephanorhynchus White, 1846) found in these areas [18]. The description of this exclusively alpine weevil genus increases our understanding of the diversity of Coleoptera found in these habitats.
High-altitude eugnomine weevil richness and abundance may correlate with the high diversity of alpine plants. There are approximately 600 species of plants in the Southern Alps [38]. The New Zealand alpine flora has an extraordinarily high proportion of white flowers, with the proportion of white flowered species in New Zealand being double that compared of other alpine regions in the world [39]. It is thought that the pollinator fauna is rather depauperate and unspecialised [40], but the presence of eugnomine weevils and other beetles found exclusively on alpine flowers indicates that there is some level of host plant specialisation [41,42]. Although beetles have been frequently acknowledged as being frequent flower visitors [39,40,43,44], they have not yet received specific attention to evaluate their role or effectiveness as pollinators in New Zealand ecosystems. Research in other countries has revealed a number of systems in which beetles, and weevils specifically, are primarily pollinators [45,46,47,48,49]. It is possible that Kuschelysius and other alpine weevils may play an important role in pollination of alpine plants, including the pollination of high-altitude populations of the forest-inhabiting Dracophyllum traversii.
The difference in wing size between K. hollowayae and K. durus suggests that the importance of flight is very different for these two taxa. The shrubland and alpine forest habitat where K. hollowayae is found, is likely to require greater flight abilities than the tussock grassland and herbfields inhabited by K. durus, consistent with hypotheses that more homogenous environments promote flightlessness [50]. Of these two, the larger species, K. hollowayae, was fully winged, the opposite of the general trend in New Zealand alpine stoneflies [51]. Wing reduction or loss occurs as part of a syndrome of characters that repeatedly evolve in alpine insect taxa [52,53], like having a dark pigmented cuticle [37], or presence of quiescence instead of diapause [36]. Flightlessness caused by wing reduction is likely to be one reason driving the species diversification in New Zealand alpine environments. In the Lucanidae, for example, wing reduction seems to have promoted speciation through the isolation of localised allopatric populations [54].
A full appreciation of the evolution of New Zealand’s eugnomine weevil fauna, its relationships to host plants and the origin of the alpine fauna will require phylogenetic and faunistic studies coupled with more natural history observations. Furthermore, we expect to find additional species of Kuschelysius, especially in alpine areas with limited access. This paper is the first installment of what we hope will be a series of papers describing the New Zealand fauna and forming the basis for a full systematic treatment of the Eugnomini, which will underpin ecological and evolutionary studies.
Author Contributions
S.D.J.B. and R.A.B.L. wrote the paper. S.D.J.B. prepared the illustrations.
Funding
This research was supported by the Strategic Science Investment Funding from the Ministry of Business, Innovation and Employment’s Science and Innovation Group to Plant and Food Research (S.D.J.B.) and to Manaaki Whenua Landcare Research (R.A.B.L.).
Acknowledgments
Birgit Rhode took habitus photographs. Dalin Brown and Rolf Oberprieler helped with suggesting and clarifying name formation. David Logan and John Marris provided valuable feedback on the manuscript. The base layer of the distribution map was provided courtesy of Geographx.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Caldara, R.; Franz, N.M.; Oberprieler, R.G. 3.7.10 Curculioninae Latreille, 1802. In Handbook of Zoology, Arthropoda: Insecta. Coleoptera, Beetles Volume 3: Morphology and Systematics (Phytophaga); Leschen, R.A.B., Beutel, R.G., Eds.; Walter De Gruyter: Berlin, Germany, 2014; pp. 589–628. [Google Scholar]
- Mazur, M.A. Koghicola karolinae, new genus and species of the Eugnomini from New Caledonia (Coleoptera: Curculionidae). Ann. Zool. 2014, 64, 9–14. [Google Scholar] [CrossRef]
- Cawthra, E.M. A redefinition of the subfamily Eugnominae to include the Meriphinae. Trans. R. Soc. N. Z. Zool. 1966, 7, 171–178. [Google Scholar]
- May, B.M. Larvae of Curculionoidea (Insecta: Coleoptera): A systematic overview. Fauna N. Z. 1993, 28, 1–223. [Google Scholar]
- Mazur, M.A. First record of the tribe Eugnomini Lacordaire, 1863 (Coleoptera: Curculionidae) from Fiji with description of Pactola fiji sp. n. Zootaxa 2012, 3517, 63–70. [Google Scholar]
- Mazur, M.A. Review of the New Caledonian species of the genus Pactola Pascoe, 1876 (Coleoptera: Curculionidae: Eugnomini), with description of two new species. Zootaxa 2014, 3814, 202–220. [Google Scholar] [CrossRef] [PubMed]
- Mazur, M.A. A new genus and nine new species of Eugnomini (Coleoptera, Curculionidae) from New Caledonia. ZooKeys 2016, 554, 87–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazur, M.A.; Tron, F.; Mille, C. Pactola kuscheli sp. nov. (Coleoptera: Curculionidae), a potential cause for the decline of the threatened New Caledonian conifer, Agathis montana de Laubenfels, 1969 (Araucariaceae). Austral Entomol. 2017, 56, 268–276. [Google Scholar] [CrossRef]
- Mazur, M.A.; Jezuita, A. A new species of Pactola Pascoe, 1876 (Coleoptera: Curculionidae: Eugnomini) from New Caledonia. Zootaxa 2015, 3931, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Broun, T. Manual of the New Zealand Coleoptera; Government Printer: Wellington, New Zealand, 1880; Volume 1.
- Broun, T. Manual of the New Zealand Coleoptera; Government Printer: Wellington, New Zealand, 1893; Volumes 5–7.
- Broun, T. Manual of the New Zealand Coleoptera; Government Printer: Wellington, New Zealand, 1886; Volumes 3 and 4.
- Broun, T. Descriptions of new genera and species of Coleoptera. Bull. N. Z. Inst. 1914, 1, 143–266. [Google Scholar] [CrossRef]
- Broun, T. Descriptions of new genera and species of Coleoptera. Trans. Proc. N. Z. Inst. 1913, 45, 97–163. [Google Scholar]
- Hudson, G.V. Fragments of New Zealand Entomology; Ferguson & Osborn Ltd.: Wellington, New Zealand, 1950. [Google Scholar]
- Hudson, G.V. New Zealand Beetles and Their Larvae: An Elementary Introduction to the Study of Our Native Coleoptera; Ferguson & Osborn Ltd.: Wellington, New Zealand, 1934. [Google Scholar]
- Marshall, G.A.K. New Curculionidae (Col.) from New Zealand. Trans. R. Soc. N. Z. 1937, 67, 316–340. [Google Scholar]
- Brown, S.D.J. Austromonticola, a new genus of broad-nosed weevil (Coleoptera: Curculionidae: Entiminae) from montane areas of New Zealand. ZooKeys 2017, 707, 73–130. [Google Scholar] [CrossRef] [PubMed]
- Kuschel, G. New Zealand Curculionoidea from Captain Cook’s voyages (Coleoptera). N. Z. J. Sci. 1970, 13, 191–205. [Google Scholar]
- Kuschel, G. Insects of Campbell Island. Coleoptera: Curculionidae of the subantarctic islands of New Zealand. Pac. Isl. Monogr. 1964, 7, 415–493. [Google Scholar]
- Kuschel, G. Apionidae and Curculionidae (Coleoptera) from the Poor Knights Islands, New Zealand. J. R. Soc. N. Z. 1982, 12, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Kuschel, G. Beetles in a Suburban Environment: A New Zealand Case Study; DSIR Plant Protection Report Volume 3; Department of Scientific and Industrial Research: Auckland, New Zealand, 1990. [Google Scholar]
- Oberprieler, R.G.; Anderson, R.S.; Marvaldi, A.E. 3 Curculionoidea Latreille, 1802: Introduction, Phylogeny. In Handbook of Zoology, Arthropoda: Insecta. Coleoptera, Beetles Volume 3: Morphology and Systematics (Phytophaga); Leschen, R.A.B., Beutel, R.G., Eds.; Walter De Gruyter: Berlin, Germany, 2014; pp. 285–300. [Google Scholar] [CrossRef]
- Lawrence, J.F.; Beutel, R.G.; Leschen, R.A.B.; Ślipiński, A. 2. Glossary of Morphological Terms. In Handbuch der Zoologie/Handbook of Zoology. Band/Volume IV Arthropoda: Insecta Teilband/Part 38. Coleoptera, Beetles. Vol. 2. Morphology and Systematics (Polyphaga Partim); Leschen, R.A.B., Beutel, R.G., Lawrence, J.F., Eds.; W. DeGruyter: Berlin, Germany, 2010; pp. 9–20. [Google Scholar] [CrossRef]
- Wanat, M. Alignment and homology of male terminalia in Curculionoidea and other Coleoptera. Invertebr. Syst. 2007, 21, 147–171. [Google Scholar] [CrossRef]
- Kukalová-Peck, J.; Lawrence, J.F. Evolution of the hind wing in Coleoptera. Can. Entomol. 1993, 125, 181–258. [Google Scholar] [CrossRef]
- Zherikhin, V.V.; Gratshev, V.G. A comparative study of the hind wing venation of the superfamily Curculionoidea, with phylogenetic implications. In Biology, Phylogeny and Classification of Coleoptera. Papers Celebrating the 80th Birthday of Roy A Crowson; Pakaluk, J., Slipinski, S.A., Eds.; Muzeum i Instytut Zoologii PAN: Warszawa, Poland, 1995; pp. 633–777. [Google Scholar]
- Kelly, K.L.; Judd, D.B. Color: Universal Language and Dictionary of Names; U.S. Department of Commerce, National Bureau of Standards: Washington, DC, USA, 1976.
- Jaffer, A. Color-Name Dictionaries: NBS/ISCC centroids. Available online: http://people.csail.mit.edu/jaffer/Color/Dictionaries#nbs-iscc (accessed on 22 July 2018).
- Inkscape Team. Inkscape: A Vector Drawing Tool. Available online: http://www.inkscape.org (accessed on 22 July 2018).
- Crosby, T.K.; Dugdale, J.S.; Watt, J.C. Area codes for recording specimen localities in the New Zealand subregion. N. Z. J. Zool. 1998, 25, 175–183. [Google Scholar] [CrossRef] [Green Version]
- Brockhampton Press. Dictionary of Classical Mythology; Brockhampton Press: London, UK, 1995. [Google Scholar]
- Hoare, R.J.B. A new species of Hierodoris Meyrick (Lepidoptera: Oecophoridae) with a telescopic ovipositor, from granite sand plains in Fiordland. N. Z. Entomol. 2012, 35, 51–57. [Google Scholar] [CrossRef]
- Hoare, R.J.B.; Millar, I.; Richardson, S.J. The insect fauna of granite sand plains: A naturally rare ecosystem in New Zealand. N. Z. Entomol. 2016, 39, 1–14. [Google Scholar] [CrossRef]
- Barratt, B.I.P.; Patrick, B.H. Insects of snow tussock grassland on the East Otago Plateau. N. Z. Entomol. 1987, 10, 69–98. [Google Scholar] [CrossRef]
- Buckley, T.R.; Krosch, M.; Leschen, R.A.B. Evolution of New Zealand insects: Summary and prospectus for future research. Austral Entomol. 2015, 54, 1–27. [Google Scholar] [CrossRef]
- Leschen, R.A.B.; Buckley, T.R. Revision and phylogeny of Syrphetodes (Coleoptera: Ulodidae): implications for biogeography, alpinization and conservation. Syst. Entomol. 2015, 40, 143–168. [Google Scholar] [CrossRef]
- Heenan, P.B.; McGlone, M.S. Evolution of New Zealand alpine and open-habitat plant species during the late Cenozoic. N. Z. J. Ecol. 2013, 37, 105–113. [Google Scholar]
- Wardle, P. Origin of the New Zealand mountain flora, with special reference to trans-Tasman relationships. N. Z. J. Bot. 1978, 16, 535–550. [Google Scholar] [CrossRef] [Green Version]
- Primack, R.B. Insect pollination in the New Zealand mountain flora. N. Z. J. Bot. 1983, 21, 317–333. [Google Scholar] [CrossRef] [Green Version]
- Webb, C.J.; Kelly, D. The reproductive biology of the New Zealand flora. Trends Ecol. Evol. 1993, 8, 442–447. [Google Scholar] [CrossRef]
- Minoshima, Y.N.; Seidel, M.; Wood, J.R.; Leschen, R.A.B.; Gunter, N.L.; Fikáček, M. Morphology and biology of the flower-visiting water scavenger beetle genus Rygmodus (Coleoptera: Hydrophilidae). Entomol. Sci. 2018. [Google Scholar] [CrossRef]
- Godley, E.J. Flower biology in New Zealand. N. Z. J. Bot. 1979, 17, 441–466. [Google Scholar] [CrossRef] [Green Version]
- Newstrom, L.; Robertson, A.W. Progress in understanding pollination systems in New Zealand. N. Z. J. Bot. 2005, 43, 1–59. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, J.E.; Irvine, A.K. Floral biology of Myristica insipida (Myristicaceae), a distinctive beetle pollination syndrome. Am. J. Bot. 1989, 76, 86–94. [Google Scholar] [CrossRef]
- Pellmyr, O.; Thien, L.B.; Bergström, G.; Groth, I. Pollination of New Caledonian Winteraceae: Opportunistic shifts or parallel radiation with their pollinators? Plant Syst. Evol. 1990, 173, 143–157. [Google Scholar] [CrossRef]
- Hall, J.A.; Walter, G.H.; Bergstrom, D.M.; Machin, P.J. Pollination ecology of the Australian cycad Lepidozamia peroffzkyana (Zamiaceae). Aust. J. Bot. 2004, 52, 333–343. [Google Scholar] [CrossRef]
- Ratnayake, R.M.C.S.; Gunatilleke, I.A.U.N.; Wijesundara, D.S.A.; Saunders, R.M.K. Pollination ecology and breeding system of Xylopia championii (Annonaceae): Curculionid beetle pollination, promoted by floral scents and elevated floral temperatures. Int. J. Plant Sci. 2007, 168, 1255–1268. [Google Scholar] [CrossRef] [Green Version]
- Franz, N.M. Pollination of Anthurium (Araceae) by derelomine flower weevils (Coleoptera: Curculionidae). Int. J. Trop. Biol. 2007, 55, 269–277. [Google Scholar] [CrossRef]
- Roff, D.A. The evolution of flightlessness in insects. Ecol. Monogr. 1990, 60, 389–421. [Google Scholar] [CrossRef]
- McCulloch, G.A.; Waters, J.M. Does wing reduction influence the relationship between altitude and insect body size? A case study using New Zealand’s diverse stonefly fauna. Ecol. Evol. 2018, 8, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Darlington, P.J. Carabidae of mountains and islands: Data on the evolution of isolated faunas, and on atrophy of wings. Ecol. Monogr. 1943, 13, 37–61. [Google Scholar] [CrossRef]
- Mani, M.S. Ecology and Biogeography of High Altitude Insects; Dr W. Junk Publishers: The Hague, The Netherlands, 1968. [Google Scholar] [CrossRef]
- Holloway, B.A. Wing development and evolution of New Zealand Lucanidae (Insecta: Coleoptera). Trans. R. Soc. N. Z. 1963, 3, 99–116. [Google Scholar]
Figure 1.
Distribution of Kuschelysius hollowayae (circles), K. verbalis (square), K. nitens (triangle) and K. durus (stars) in the South Island of New Zealand. Two-letter codes indicate the regions defined by Crosby et al. [31].
Figure 1.
Distribution of Kuschelysius hollowayae (circles), K. verbalis (square), K. nitens (triangle) and K. durus (stars) in the South Island of New Zealand. Two-letter codes indicate the regions defined by Crosby et al. [31].

Figure 2.
Habitus figures of Kuschelysius species. (a) K. hollowayae, dorsal view; (b) K. hollowayae, lateral view; (c) K. durus, dorsal view; (d) K. durus, lateral view; (e) K. verbalis holotype, dorsal view; (f) K. verbalis holotype, lateral view; (g) K. nitens holotype, dorsal view; (h) K. nitens holotype, lateral view. Scale bars = 1 mm.
Figure 2.
Habitus figures of Kuschelysius species. (a) K. hollowayae, dorsal view; (b) K. hollowayae, lateral view; (c) K. durus, dorsal view; (d) K. durus, lateral view; (e) K. verbalis holotype, dorsal view; (f) K. verbalis holotype, lateral view; (g) K. nitens holotype, dorsal view; (h) K. nitens holotype, lateral view. Scale bars = 1 mm.

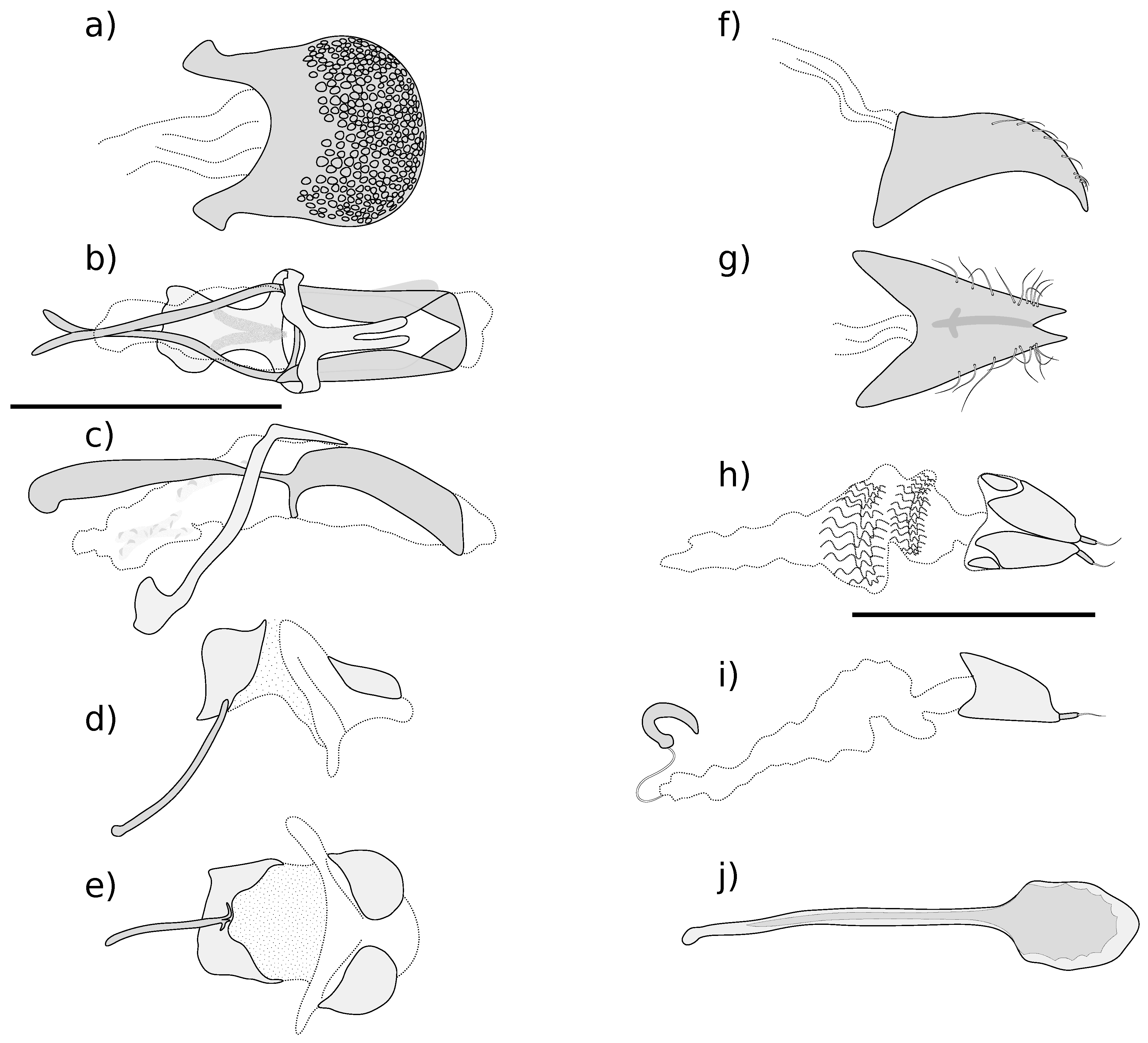
Figure 3.
Hind wing of Kuschelysius species. (a) K. hollowayae; (b) K. durus. AA: anal anterior vein; ac: anal cell; AP: anal posterior vein; C: costal vein; CuA: cubitus anterior vein; J: jugal vein; MP: media posterior veins; ms: medial spur; msc: medial sclerotisation; RA: radius anterior vein; RC: radial cell; rf: radial fold; RP: radius posterior veins; rs: radial sclerites; Sc: Subcostal vein. Wings drawn to the same scale; scale bar = 1 mm.
Figure 3.
Hind wing of Kuschelysius species. (a) K. hollowayae; (b) K. durus. AA: anal anterior vein; ac: anal cell; AP: anal posterior vein; C: costal vein; CuA: cubitus anterior vein; J: jugal vein; MP: media posterior veins; ms: medial spur; msc: medial sclerotisation; RA: radius anterior vein; RC: radial cell; rf: radial fold; RP: radius posterior veins; rs: radial sclerites; Sc: Subcostal vein. Wings drawn to the same scale; scale bar = 1 mm.

Figure 4.
Right antennae, anterior view, of Kuschelysius species. (a) K. hollowayae; (b) K. durus; (c) K. verbalis; (d) K. nitens; scale bar = 1 mm.
Figure 4.
Right antennae, anterior view, of Kuschelysius species. (a) K. hollowayae; (b) K. durus; (c) K. verbalis; (d) K. nitens; scale bar = 1 mm.

Figure 5.
Left femora and tibae, anterior view, of Kuschelysius species. Kuschelysius hollowayae: (a) profemur and tibia; (b) mesofemur and tibia; (c) metafemur and tibia; Kuschelysius durus: (d) profemur and tibia; (e) mesofemur and tibia; (f) metafemur and tibia; Kuschelysius verbalis: (g) profemur and tibia; (h) mesofemur and tibia; (i) metafemur and tibia; Kuschelysius nitens: (j) profemur and tibia; (k) mesofemur and tibia; (l) metafemur and tibia; scale bar = 1 mm.
Figure 5.
Left femora and tibae, anterior view, of Kuschelysius species. Kuschelysius hollowayae: (a) profemur and tibia; (b) mesofemur and tibia; (c) metafemur and tibia; Kuschelysius durus: (d) profemur and tibia; (e) mesofemur and tibia; (f) metafemur and tibia; Kuschelysius verbalis: (g) profemur and tibia; (h) mesofemur and tibia; (i) metafemur and tibia; Kuschelysius nitens: (j) profemur and tibia; (k) mesofemur and tibia; (l) metafemur and tibia; scale bar = 1 mm.

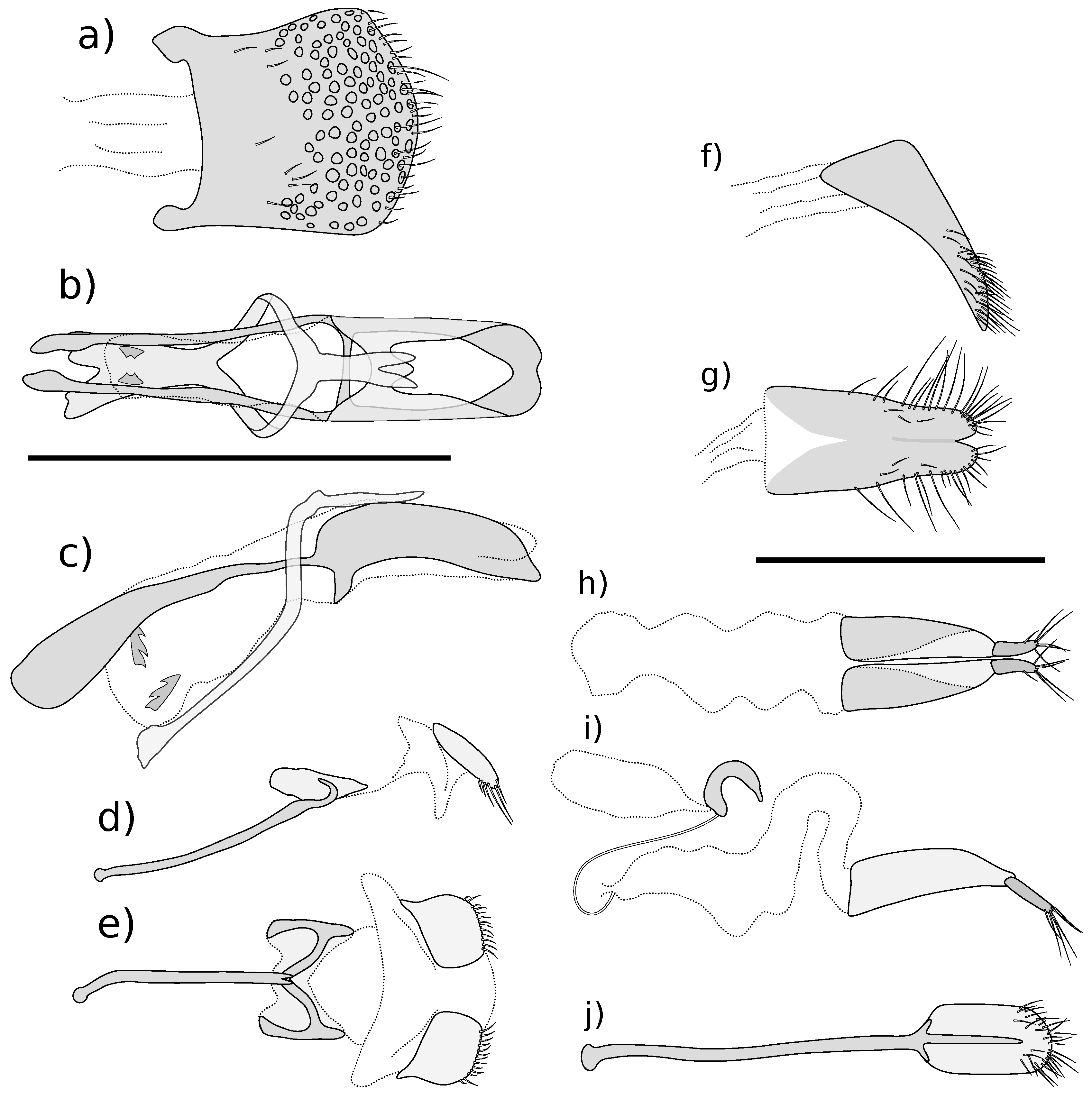
Figure 6.
Male and female genitalia of Kuschelysius hollowayae. (a) male tergite 8, dorsal view; (b) aedeagus, dorsal view; (c) aedeagus, lateral view; (d) male sternite 8, lateral view; (e) male sternite 8, ventral view; (f) female tergite 8, lateral view; (g) female tergite 8, dorsal view; (h) gonocoxites and bursa copulatrix, dorsal view; (i) gonocoxites, bursa copulatrix and spermatheca, lateral view; (j) female sternite 8, ventral view. Scale bars = 1 mm, a–e at same scale, f–j at same scale.
Figure 6.
Male and female genitalia of Kuschelysius hollowayae. (a) male tergite 8, dorsal view; (b) aedeagus, dorsal view; (c) aedeagus, lateral view; (d) male sternite 8, lateral view; (e) male sternite 8, ventral view; (f) female tergite 8, lateral view; (g) female tergite 8, dorsal view; (h) gonocoxites and bursa copulatrix, dorsal view; (i) gonocoxites, bursa copulatrix and spermatheca, lateral view; (j) female sternite 8, ventral view. Scale bars = 1 mm, a–e at same scale, f–j at same scale.

Figure 7.
Male and female genitalia of Kuschelysius durus. (a) male tergite 8, dorsal view; (b) aedeagus, dorsal view; (c) aedeagus, lateral view; (d) male sternite 8, lateral view; (e) male sternite 8, ventral view; (f) female tergite 8, lateral view; (g) female tergite 8, dorsal view; (h) gonocoxites and bursa copulatrix, dorsal view; (i) gonocoxites, bursa copulatrix and spermatheca, lateral view; (j) female sternite 8, ventral view. Scale bars = 1 mm, (a–e) at same scale, (f–j) at same scale.
Figure 7.
Male and female genitalia of Kuschelysius durus. (a) male tergite 8, dorsal view; (b) aedeagus, dorsal view; (c) aedeagus, lateral view; (d) male sternite 8, lateral view; (e) male sternite 8, ventral view; (f) female tergite 8, lateral view; (g) female tergite 8, dorsal view; (h) gonocoxites and bursa copulatrix, dorsal view; (i) gonocoxites, bursa copulatrix and spermatheca, lateral view; (j) female sternite 8, ventral view. Scale bars = 1 mm, (a–e) at same scale, (f–j) at same scale.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Brown, S.D.J.; Leschen, R.A.B. Kuschelysius, a New Alpine Genus of Eugnomine Weevil (Coleoptera: Curculionidae: Curculioninae) from New Zealand. Diversity 2018, 10, 75. https://doi.org/10.3390/d10030075
AMA Style
Brown SDJ, Leschen RAB. Kuschelysius, a New Alpine Genus of Eugnomine Weevil (Coleoptera: Curculionidae: Curculioninae) from New Zealand. Diversity. 2018; 10(3):75. https://doi.org/10.3390/d10030075
Chicago/Turabian StyleBrown, Samuel D. J., and Richard A. B. Leschen. 2018. "Kuschelysius, a New Alpine Genus of Eugnomine Weevil (Coleoptera: Curculionidae: Curculioninae) from New Zealand" Diversity 10, no. 3: 75. https://doi.org/10.3390/d10030075
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.