1. Introduction
Marine coastal areas and continental shelves are generally characterized by highly productive ecosystems with elevated levels of biodiversity. The Mediterranean Sea, despite its relatively small dimensions [
1], is known as a hotspot of biodiversity [
2,
3]. In its shallow waters several biogenic structures (bioconstructions) play an important role in the high level of biodiversity, even though they are less known than those flourishing in tropical seas [
4].
The bioconstructions built by gastropods belonging to the genus
Dendropoma (Vermetidae) are commonly distributed along the shallow warm-water coasts of the Mediterranean Sea and are particularly important as biological markers of sea level fluctuations, they protect the underlying rock from the coastal erosion processes and increase the hard-bottom complexity [
5,
6,
7].
The sessile and gregarious habits of vermetids along with their capability of secreting calcium carbonate are the main features through which the biogenic structures are built up along shallow rocky coasts; these concretions attract and become colonized by a particular associated community [
8,
9,
10]. The vermetid shells overgrow each other, starting from the hard substrate on which the species primarily settled. The result is a complex bioconstruction that hosts a large number of small invertebrates, enhancing local density and species richness. The vermetid shells are structural components of habitats (
sensu Jones [
11]) and play a multiple functional role: (i) providing protection to the organisms against the physical disturbance caused by water movement and the physiological stress caused by desiccation during low tide; (ii) constituting a suitable substrate for colonization as well as a refuge from predation; (iii) supporting high resources availability at different trophic levels [
12,
13].
Current research works are mainly focused on the reproductive biology of vermetid species from different areas along the Mediterranean coasts (e.g., Spain [
14], Italy [
15], Israel [
16,
17], western and central Mediterranean [
18]). Conversely, there are few studies regarding fauna associated with the vermetid bioconstructions. In the eastern Mediterranean, along the Israeli shore, the polychaete assemblage living among vermetid shells was composed of 70 species [
13], while fish biodiversity was represented by 36 species [
19]. In the central Mediterranean, along the Sicilian shore, the algal community was represented by more than 100 species [
20], fish biodiversity by 39 taxa belonging to 15 families [
21] and the mollusc community by 46 taxa [
22] and by 28 gastropod species [
10]. Along the Western Mediterranean coast, the fauna associated with these biogenic structures is still poorly documented [
23], while recent research deals mainly with algal assemblages, represented by over 100 taxa [
24,
25].
The main purpose of the present study is therefore to investigate the role of the vermetid bioconstructions, formed by the endemic prosobranch Dendropoma lebeche Templado, Richter and Calvo 2016, in modeling the structural and functional (trophic) diversity of the associated mollusc assemblage along the Spanish Mediterranean coast.
3. Results
3.1. Composition of Mollusc Assemblages
A total of 600 individuals represented by 57 species and 31 families associated with vermetid bioconstructions were recorded. They belong to the classes of Polyplacophora (three families; four species; 37 individuals), Gastropoda (18 families; 35 species; 288 individuals) and Bivalvia (10 families; 18 species; 275 individuals) (
Table 2). These three taxa were mainly represented respectively by the families Acanthochitonidae, Rissoidae and Mytilidae, which displayed the highest number of species, while 16 families were just represented by a single species.
At each site there was one taxonomic group that was substantially more species rich or more abundant than the other taxonomic groups. At CP, gastropods were dominant both in species richness (%DQ 74.07%, 20 sp.) and abundance (%DI 72.09%, 124 ind.), followed by bivalves (%DQ 14.81%, four spp.; %DI 13.37%, 23 ind.) and polyplacophorans (%DQ 11.11%, three spp.; %DI 14.53%, 25 ind.). The most abundant and frequent species, belonging to these three taxa, were respectively the gastropod Tricolia tingitana (%DI 20.93%; %F 100%), the bivalve Mytilaster solidus (%DI 9.30%; %F 100%) and the polyplacophoran Lepidochitona sp. (%DI 4.07%; %F 100%).
At PP, gastropods and bivalves showed the same species richness (%DQ 45.45%, 10 sp.) with an abundance of 76.19% (64 ind.) and 15.47% (13 ind.) respectively, followed by polyplacophorans (%DQ 9.09%, two spp.; %DI 8.33%, seven ind.). The most abundant and frequent species were respectively the gastropod Crisilla semistriata (%DI 50%; %F 66.6%), the bivalve M. solidus (%DI 3.57%; %F 100%) and the polyplacophoran Acanthochitona fascicularis (%DI 4.76%; %F 66.6%).
At Ja, gastropods were dominant in species richness (%DQ 51.28%, 20 sp.), with an abundance of 29.06% (100 ind.), followed by bivalves (%DQ 41.02%, 16 sp.), with an abundance of 69.47% (239 ind.), while polyplacophorans were poorly represented (%DQ 7.69%, three spp.; %DI 1.45%, five ind.). The most abundant and frequent species respectively for the three taxa were the gastropod Gibbula drepanensis (%DI 7.56%; %F 100%), the bivalve M. solidus (%DI 43.60%; %F 100%) and the polyplacophoran A. fascicularis (%DI 0.58%; %F 66.6%).
3.2. Trophic Diversity
The mollusc assemblage was dominated for 51% by herbivores (MG 155 ind.; 20 spp. and DF 151 ind.; nine spp.) and 45.83% by omnivores (SF 275 ind.; 18 spp.), followed for 3.17% by carnivores (P 14 ind.; nine spp. and E five ind.; one sp.).
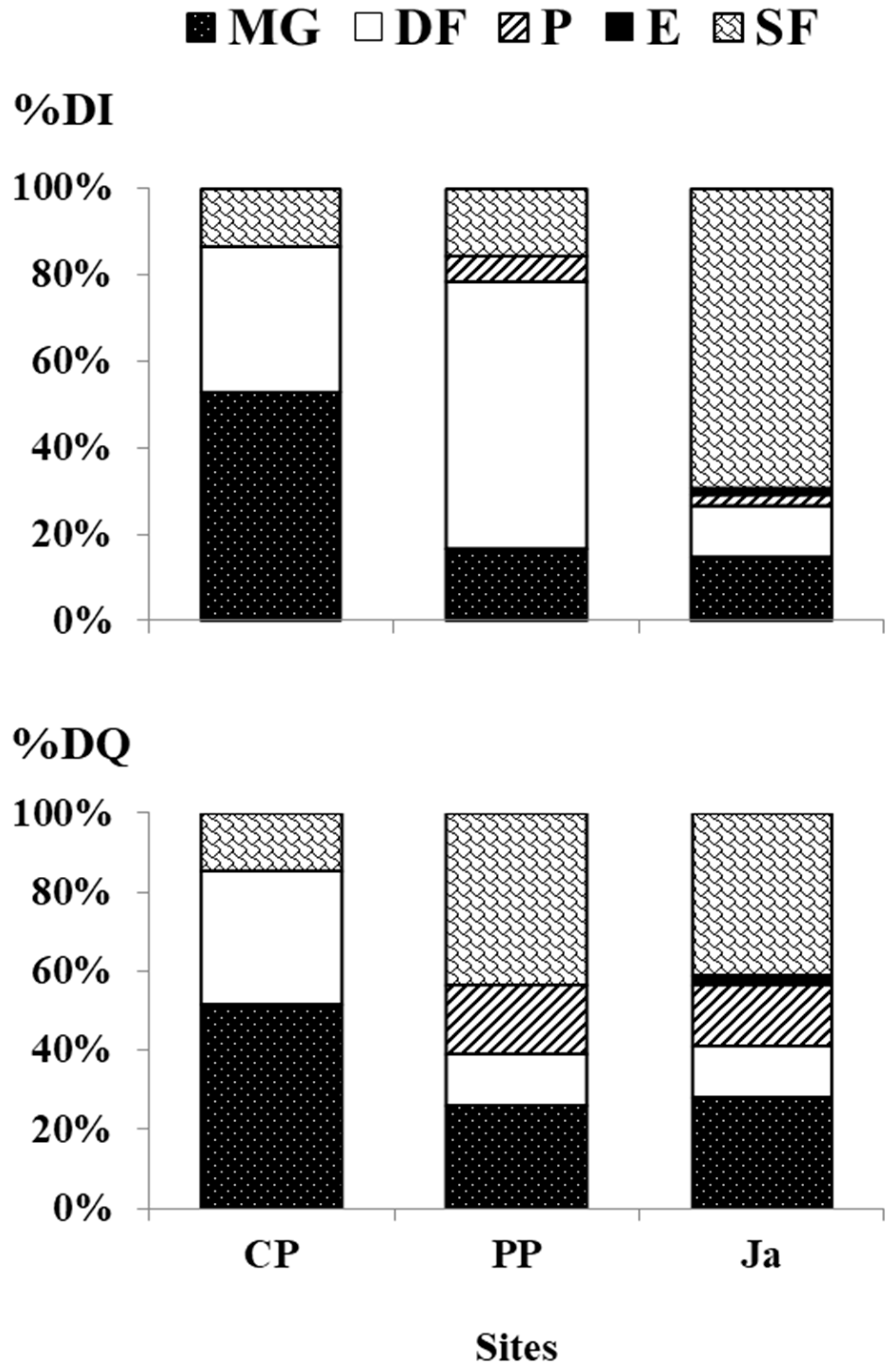
At each site, a single feeding guild showed a higher dominance upon the others (
Figure 2). At CP, the feeding guild MG was dominant both in abundance and species richness (%DI 52.91%, 91 ind.; %DQ 51.85%, 14 spp.) and was mainly represented by the gastropod
T. tingitana (36 ind.). At PP, the feeding guild DF was dominant in abundance (%DI 61.90%; 52 ind.), while SF was dominant in species richness (%DQ 43.48%; 10 spp.). These guilds were mainly represented by the gastropod
C. semistriata (42 ind.) and the bivalve
M. solidus (three ind.) respectively. At Ja, the dominant feeding guild was SF, both in abundance and species richness (%DI 69.48%, 239 ind.; %DQ 41.03%, 16 spp.) and it was mainly represented by the bivalve
M. solidus (150 ind.).
3.3. Structural Analysis of Mollusc Assemblage
A mean value of 66.66 ± 47.22 individuals/dm
2 was recorded. The maximum abundance (N) occurred at the Ja site, an intermediate value occurred at CP and the minimum value at PP. This same trend was found for species richness (SR). PERMANOVA test highlighted significant differences both for mollusc abundance and species richness among sites. Diversity indices (H’, J’), instead, resulted higher at CP than at the other two sites (
Table 3).
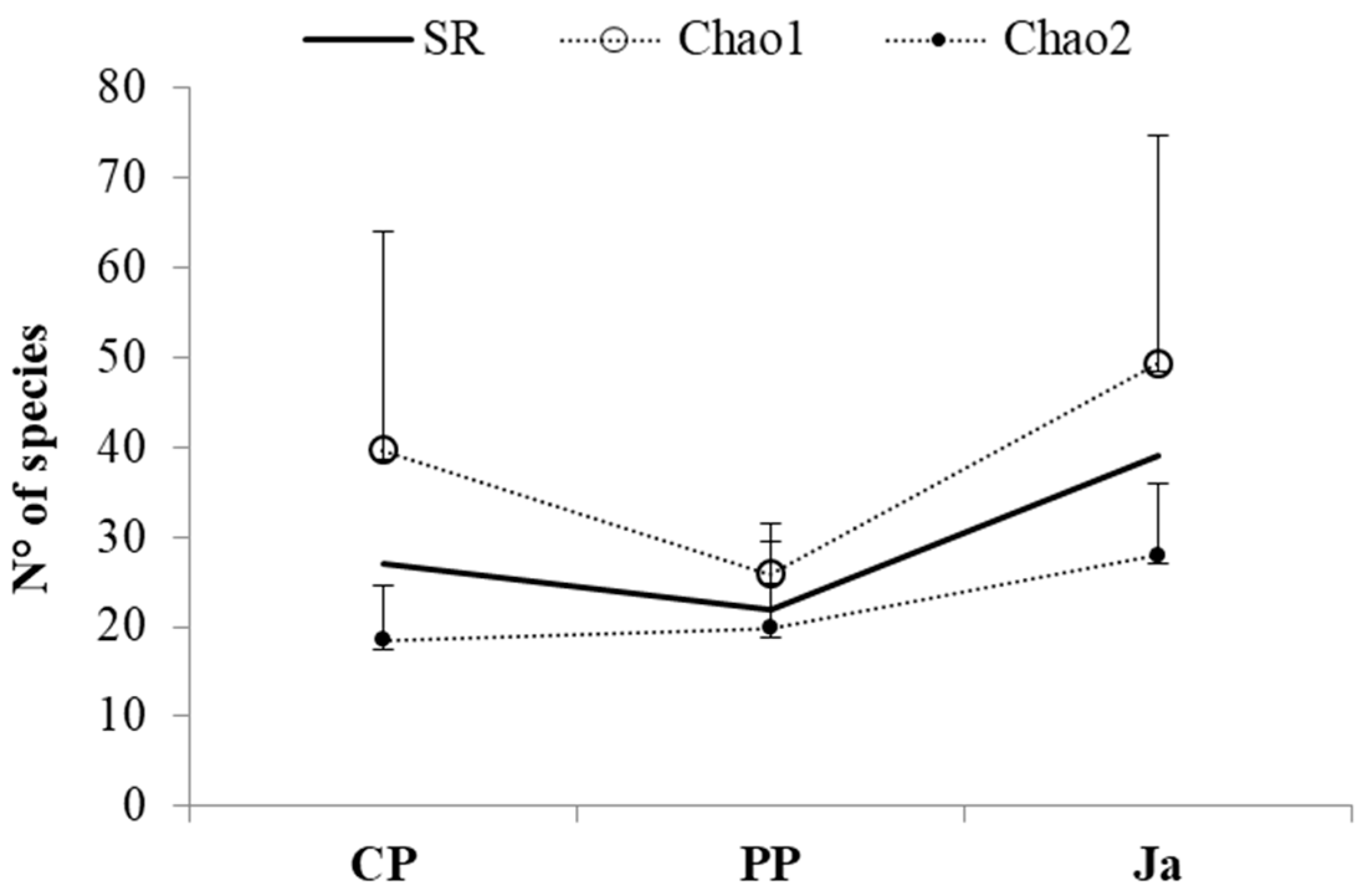
The richness estimators (Chao1 and Chao2;
Figure 3) revealed that the efficiency of sampling was higher at PP than at CP and at Ja, for both the expected number of individuals and the expected number of species.
PERMANOVA test highlighted significant differences in structure of mollusc assemblage among the three sites, with a highly significant
p-value between CP and Ja (
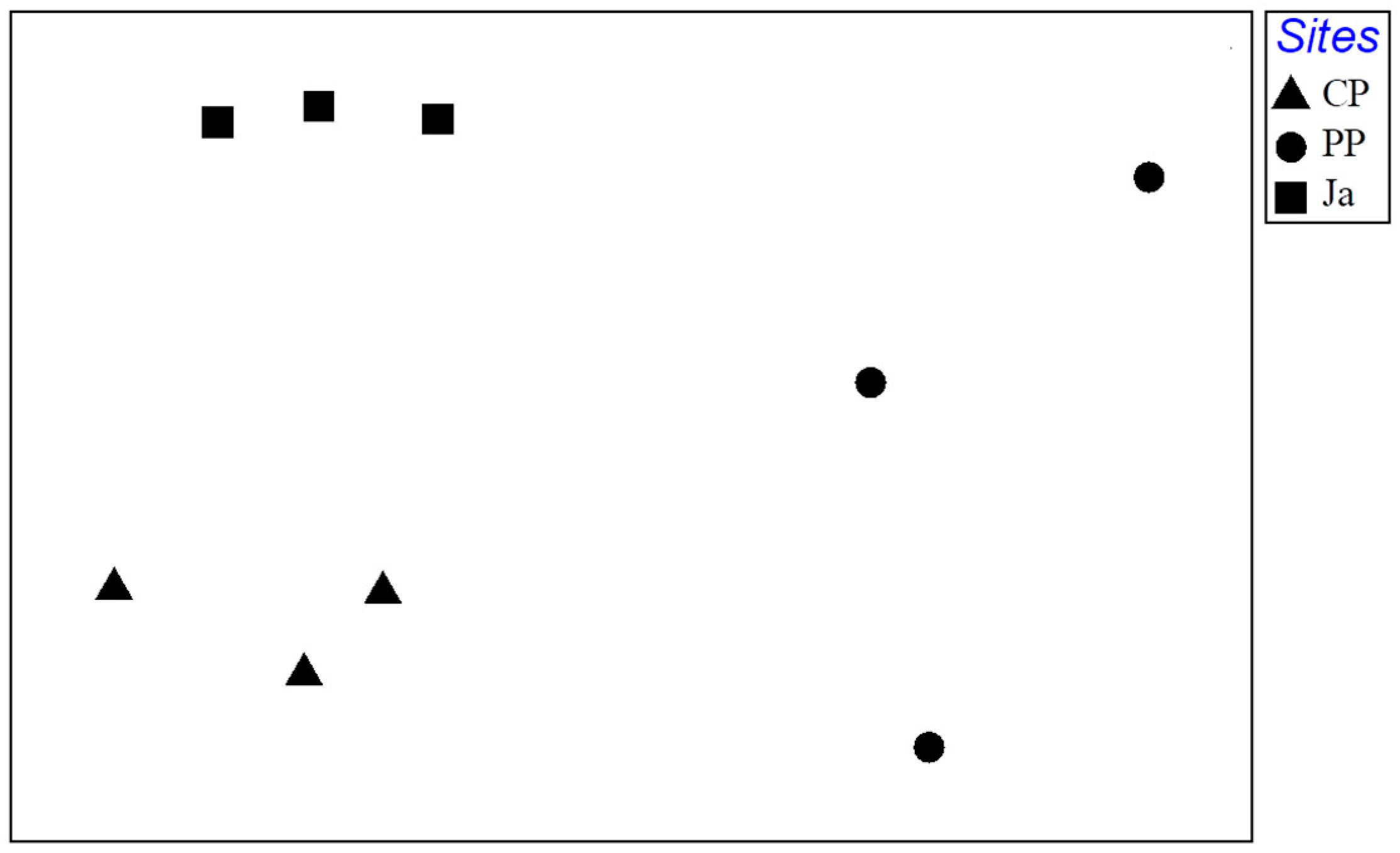
Table 4). These differences were evident in the nMDS plot (
Figure 4). CP and Ja were clearly separated and replicates formed a consistent cluster for each site, while PP showed a high dispersion of replicates between the two other sites.
SIMPER analysis, based on the mollusc composition, indicated that the highest average similarities among the replicates occurred for the sites Ja (61.88%) and CP (58.06%), while that for PP replicates was only 29.94% (
Table 5). The species that concurred for about 50% of similarity were mainly represented: at Ja by the suspension feeders, such as
M. solidus,
Rocellaria dubia,
Gregariella semigranata and
Hiatella arctica; at CP by the deposit feeder
Sinezona cingulata and grazer species
T. tingitana and
Lepidochitona sp.; at PP by the deposit feeders, such as
Setia amabilis and
Rissoa similis.
On the other hand, the average similarity among the three sites was 31.39% (
Table 6), mainly due to the suspension feeder
M. solidus and deposit feeders
R. similis,
S. cingulata and
S. amabilis.
4. Discussion
It is well known that the complex surface of vermetid bioconstructions creates semi-sheltered microhabitats, promoting an extremely diversified benthic assemblage in intertidal zone exposed to water movement. Our results showed that the mollusc assemblage occurring on the intertidal vermetid bioconstructions was composed by 600 individuals grouped in 57 species. Even though data are based on a limited number of samples, the results reflect the highest species richness ever found in vermetid bioconstructions investigations [
10,
22,
23].
Among the three taxonomic groups recorded in this study, polyplacophorans were poorly represented both in number of individuals and species richness, among which the grazers
A. fascicularis and
Acanthochitona crinita were the main representative. As highlighted in the present study, species belonging to this taxon fall into the “meso-littoral benthic component” described by Pandolfo et al. [
22] in association with the most exposed portion of Sicilian vermetid bioconstructions.
By contrast, gastropods were represented by a high abundance, species richness and trophic diversity. The grazers G. drepanensis and T. tingitana and the deposit feeders S. cingulata and C. semistriata were the most dominant herbivore species. Predators and ectoparasites were instead poorly represented, among which Marshallora adversa and Vitreolina philippi were respectively the main dominant species.
A high abundance and a rather high species richness were also detected for bivalves, such as
M. solidus,
R. dubia,
G. semigranata,
H. arctica and
Striarca lactea, even though they belong to a single trophic category represented by omnivores suspension feeders, which are relevant constituents of benthic communities of hard bottoms [
38].
The dominance of herbivores associated with bioconstruction is an indirect effect of the functional role of the vermetid ecosystem engineer in sustaining biodiversity. Previous studies [
39,
40,
41,
42] reported that the abundance and diversity of molluscan assemblages are closely related to the algal types and architecture, which represent an important food resource. This is also corroborated by Fernández et al. [
25]. They revealed a rich phytobenthic diversity inhabiting vermetid platforms of the southeastern Iberian Peninsula, with the lack of a persistent dominant group and a scarcity of grazers’ coverage where the algal cover was low. Therefore, high algal diversity growing on vermetid shells and sand coated with bacteria and organic detritus traps in the pores and crevices of bioconstruction were the main drivers of the dominant feeding guilds, such as grazers and deposit feeders.
In Sicily, Pandolfo et al. [
9] reported the bivalve
Mytilaster minimus as an important constituent of the mollusc assemblage associated with bioconstructions built up by the vermetid species
Dendropoma cristatum (Biondi 1859). Other bivalves found, such as
Lithophaga lithophaga,
Irus irus and
Petricola lithophaga, were also previously reported in association with the vermetids, both in Sicilian [
9] and Alicante coasts [
23]. These are rock-boring and filter-feeding bivalves which are especially favoured by crevice, empty vermetid shells of bioconstructions and the degree of coast exposure to the water movement. According to Österling and Pihl [
43], high water movement promotes a low algal cover that favours bivalves such as suspension feeders; while low water movement promotes a rich algal cover that favours gastropods such as grazers and their species diversity.
Even though in the three sites the vermetid bioconstructions were always sampled along the rocky shore more exposed to the water movement, different mollusc assemblages occurred and consisted of algal cover associated with the vermetid platforms reported by Fernández [
24].
Some previous studies suggested that benthic assemblages are strongly affected by environmental conditions, which changes at broader spatial scales [
44,
45,
46,
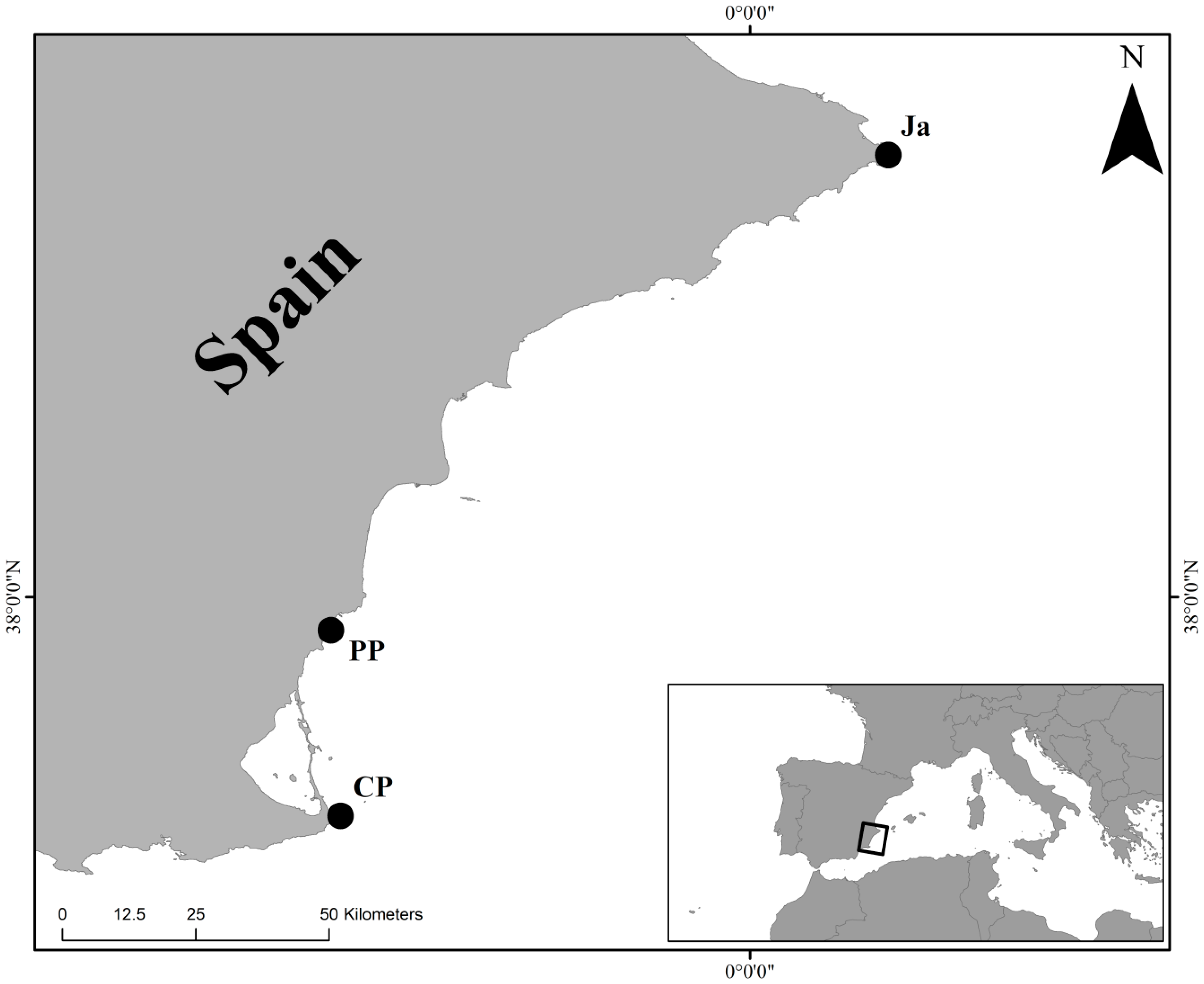
47]. Therefore, the significant difference in mollusc composition within the three sites (SIMPER, pairwise comparisons), besides bioconstructions algal covers, could be attributable to the geographical distance among the sites. Indeed, Cabo de Palos and Jávea were extremely distant sampling sites.
The westernmost site (Cabo de Palos), where vermetid bioconstructions were dominated by brown and red algae (e.g.,
Padina pavonica and
Jania sp. [
24]), was mainly characterized by herbivore gastropods including species such as
Fossarus ambiguous,
Tubbreva micrometrica,
Pisinna glabrata,
Tricolia punctura and
T. tingitana which were exclusively found there. Moreover, this latter species was reported by Gofas [
48,
49] exclusively for the coasts of southern Spain (Straits of Gibraltar area), even though it was recently detected by Scuderi and Russo [
50] along the coasts of southern Italy, both as living and fossil species.
On the other hand, the easternmost site (Jávea), where bioconstructions were dominated by red algae (e.g.,
Laurencia spp. [
24]), was mainly characterized by bivalve suspension feeders such as
Mytilus galloprovincialis,
Musculus costulatus,
R. dubia,
Arca noae and
P. lithophaga which were exclusive to this site and were previously reported for the same study area by Boronat et al. [
23].
The presence of a geographic gradient in mollusc populations, from a dominance of gastropods, at Cabo de Palos, to a dominance of bivalves, at Jávea, is confirmed by the structure of mollusc assemblage at Punta Prima. Indeed, in this latter site, located between the westernmost and the easternmost site, the mollusc assemblage was characterized by a high abundance of gastropods and high species richness of bivalves.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}